
翻译组项目 | 哪些科学问题需要从翻译调控层面寻找答案?
翻译组学作为连接转录组与蛋白质组的桥梁,通过核糖体印迹测序等核心技术(如Ribo-seq、Polysome-seq、Active Ribo-seq),揭示了生命信息传递中转录丰度与蛋白质丰度不完全等同的本质。近年来,翻译组学研究已从早期的翻译活性定量,逐步走向更深入的机制解析,重点关注翻译效率、核糖体稳态、翻译起始与延伸调控,以及 RNA 修饰等因素如何协同影响器官发育、神经功能、肿瘤代谢重塑和疾病进程【1-3】。
在许多疾病研究中,研究者常常会观察到蛋白质水平发生显著改变,但转录组数据却无法解释这一差异。此时,答案往往隐藏在翻译调控这一层。mRNA 是否真正进入翻译过程、核糖体负载是否发生变化、翻译起始和延伸是否受阻、特定密码子处是否出现核糖体停滞,以及 RNA 修饰是否影响转录本的翻译命运,共同构成了转录本到蛋白质之间一套精密的调控滤网【1-3】。
翻译组学研究的核心价值,在于回答转录组难以直接回答的问题:
这条mRNA真的被翻译了吗?翻译效率有多高?核糖体在哪个密码子处发生了停滞?是否存在 uORF、sORF、非经典开放阅读框或移码翻译等隐藏翻译事件?以 Ribo-seq 为代表的核糖体印迹测序技术,通过捕获核糖体保护片段,即 RPF,能够以近单密码子分辨率描绘特定状态下的翻译图谱,为解析翻译起始、延伸、暂停和终止等过程提供关键证据【1,4】。Active Ribo-seq 则可进一步富集活跃翻译状态下的核糖体信号,从而降低背景干扰并提高对有效翻译事件的识别能力【5】。此外,Polysome-seq 可通过多聚核糖体分级评估转录本的整体翻译活性【6】。
在医学研究中,翻译组学已经在多个前沿方向释放独特价值。
在肿瘤领域,癌细胞可通过调控特定 tRNA、氨酰 tRNA 合成酶及密码子使用偏好,形成对特定氨基酸或翻译程序的依赖【7,8】。Ribo-seq 不仅能够捕捉这类翻译重编程事件,还可用于发现 uORF、sORF、非编码区翻译和移码翻译等非经典翻译产物,为新抗原候选肽段识别、肿瘤疫苗开发和免疫治疗靶点挖掘提供重要线索【9】。近年来,靶向 tRNA 依赖性氨基酸利用的研究也进一步提示,肿瘤细胞的特定翻译依赖可能构成可干预的代谢脆弱性【10】。
在神经科学领域,突触局部翻译具有高度时空特异性,传统稳态蛋白质组学往往难以捕捉其快速变化。Ribo-seq 结合时间序列或特定神经活动模型,可帮助解析活动依赖性翻译调控,推动记忆形成、突触可塑性和神经退行性疾病机制研究;例如,近期研究显示,抑制 YTHDF2 介导的 m6A 修饰 mRNA 降解可增强神经元蛋白合成并改善海马依赖性记忆,提示 RNA 修饰、mRNA 稳定性与翻译调控之间存在紧密联系【11】。
在心血管与代谢疾病研究中,翻译组学能够在心脏发育、成熟和病理重构等关键阶段,识别代谢基因、收缩功能相关基因及应激响应基因的翻译调控节点,为心肌病和代谢异常的机制解析提供更精细的分子图谱【12】。此外,在肠道稳态研究中,Ribo-seq 也可用于区分 mRNA 丰度变化与翻译效率变化,帮助揭示核糖体稳态异常如何影响屏障功能相关蛋白的生成【13】。
在免疫与感染领域,T 细胞、巨噬细胞等免疫细胞在激活过程中会迅速发生翻译重编程,并与转录调控共同塑造细胞因子产生、代谢转换和效应功能【14】。因此,翻译组学已然是解析免疫激活早期事件、病原感染应答和炎症调控机制的重要工具。
表观生物翻译组项目已助力客户发表Nature、Cell等多篇国际顶级期刊

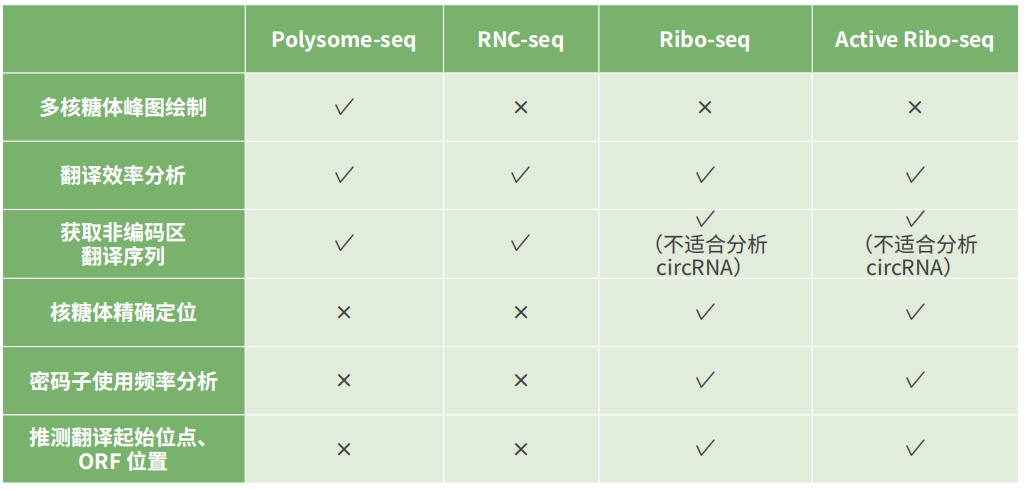
多种翻译组技术对比
开展翻译组研究前,厘清研究逻辑:以科学问题为起点,选择合适的样本和技术路线,并提前设计数据解读与功能验证方案。
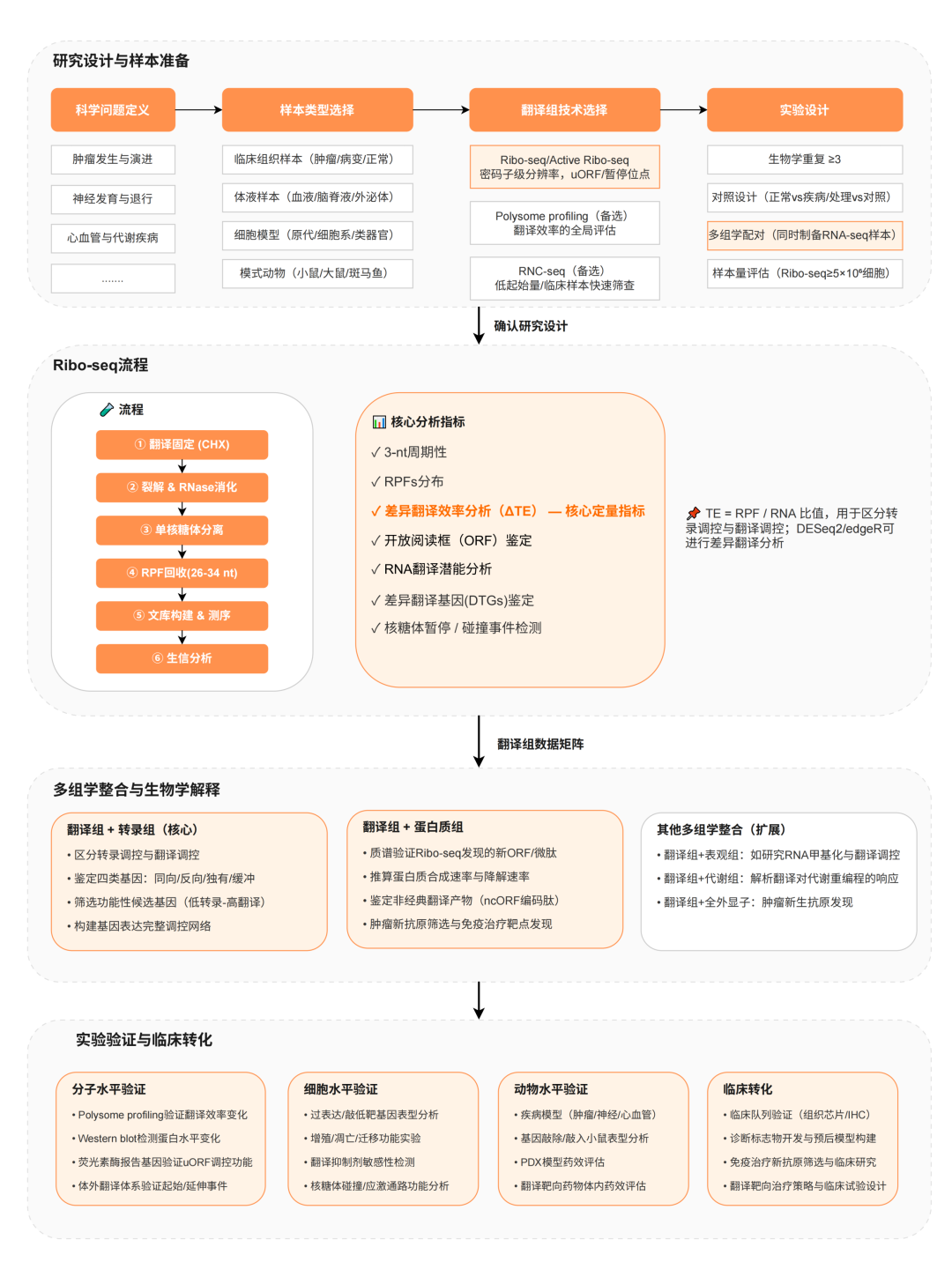
一个完整的翻译组研究框架,从科学问题出发——是肿瘤的翻译重编程、神经元的局部合成、还是心脏发育的代谢切换——然后倒推样本类型和技术选型。Ribo-seq(或Active Ribo-seq)是核心,密码子级分辨率、uORF鉴定、暂停位点检测,这些是它独有的能力;Polysome-seq做全局翻译效率的补充评估;低起始量场景下RNC-seq是临床样本快速筛查的备选方案。
数据出来之后,真正的工作才开始。ΔTE分析是翻译组的核心定量逻辑,本质上是用RPF/RNA的比值把"转录调控"和"翻译调控"拆开来看。很多研究在这一步才发现,蛋白水平的变化根本不来自mRNA的增减,而是翻译效率本身出了问题。配合蛋白质组做ncORF微肽验证,配合RNA修饰(如m6A)解析表观修饰对翻译的介入,配合代谢组追踪氨基酸通量与核糖体行为的耦合——每一层整合都在回答一个转录组单独回答不了的问题。验证链条从分子到细胞到动物模型,最终指向临床队列和治疗靶点,这是翻译组研究能走多远的关键。
那么有四篇表观生物翻译组项目近期见刊的客户文章,跟随小编看看这四篇文章的研究思路,或许能让这套逻辑更具体一些:
表观生物Ribo-seq项目案例
肠道生理

研究背景:
肠上皮细胞稳态对于维持肠屏障功能至关重要。虽然SETD2在转录调控中的作用已广为人知,但临床样本中发现SETD2缺失会导致严重的转录本与蛋白质丰度失衡,这种矛盾现象提示在后转录或翻译水平存在未知的调控环节。
研究思路与方法:
模型系统:肠上皮特异性Setd2敲除小鼠模型。利用DSS诱导急性肠损伤以评估表型。通过监测小鼠体重变化、肠道组织病理切片及肠渗透性分析,系统性评估SETD2缺失对肠屏障完整性的影响。
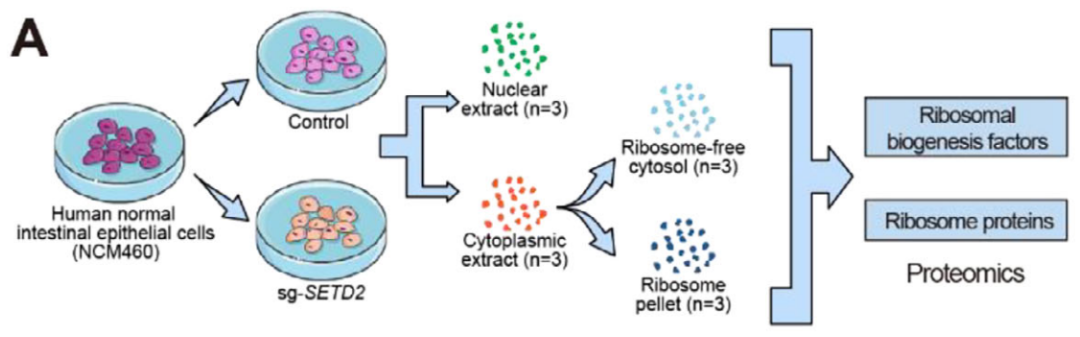
多组学集成:这篇研究整合了转录组、蛋白质组以及Ribo-seq技术,对比野生型与敲除组在损伤状态下的分子表达谱差异。通过对比mRNA丰度与实际产出的蛋白质水平,研究锁定了ANXA1与CLDN2等翻译效率下调的核心屏障基因。
机制解析:研究通过超速离心技术分离细胞核、细胞质及核糖体组分,并配合免疫共沉淀质谱分析SETD2的相互作用蛋白。利用多聚核糖体分析验证特定候选基因在各级核糖体组分中的分布,解析核糖体发生因子空间分配异常对翻译起始的阻碍。
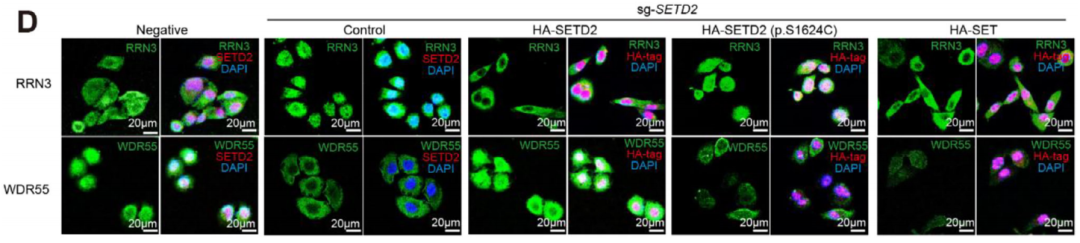
Ribo-seq的核心价值:
该研究中,多种屏障蛋白的mRNA水平并未随SETD2缺失而改变,导致传统转录组分析失效。Ribo-seq解决了由于mRNA水平波动微弱而无法解释蛋白大幅下降的困境,证明了SETD2缺失是通过干扰核糖体组分的募集,从而在特定基因的翻译起始与延伸阶段制造了障碍。
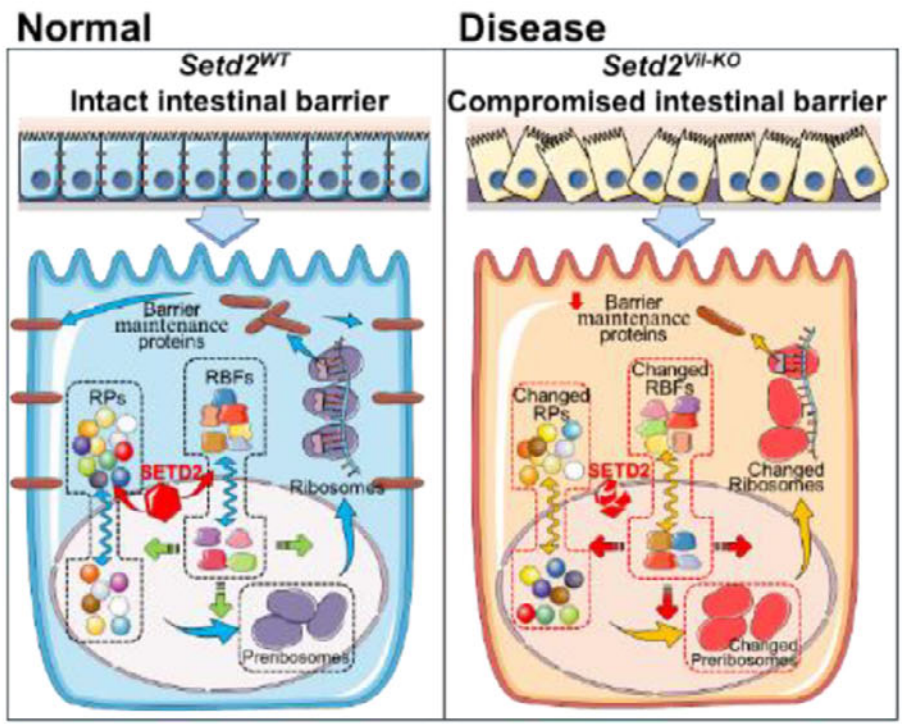
注:SETD2通过调控核糖体稳态维持肠屏障完整性
核心发现:
SETD2缺失会导致核糖体合成因子RRN3和WDR55在细胞核与细胞质间的募集异常。这种核糖体发生过程的紊乱直接下调了ANXA1、CLDN2等屏障维持基因的翻译效率,最终导致肠屏障功能的崩溃。
研究确立了SETD2在保障核糖体稳态中的关键定位,证明核糖体发生因子的空间募集障碍是导致肠屏障受损的核心翻译调控机制。
神经科学

研究背景:
m6A修饰通过调节mRNA的命运来平衡基因表达。目前该领域存在一个显著的科学悖论:通常认为阅读器蛋白YTHDF2仅负责促进mRNA降解,即清理过期转录本,但研究发现其缺失后却意外导致海马神经元内活动依赖性蛋白合成的显著增加并提升了记忆能力。该研究旨在解析阻断这种清理过程为何能反向释放神经系统的认知潜能。
研究思路与方法:
模型系统:研究先是培育了前脑特异性YTHDF2条件性敲除小鼠,在不影响大脑整体形态发育的前提下实现了目标蛋白的精准去除。通过严格的繁育策略与免疫组织化学实验验证敲除效率,确保后续行为学实验数据的特异性。
功能评估:通过恐惧调控实验和Barnes迷宫精准评估小鼠的海马依赖性记忆表现。同时应用电生理膜片钳技术记录突触后电流和长时程增强效应,从生理学角度探究YTHDF2缺失对神经元突触可塑性的支撑作用。
分子监测:团队结合RIP-seq技术鉴定YTHDF2在海马组织中直接结合的靶mRNA,并通过核糖体印迹测序监测学习行为触发后的翻译动态。应用SUnSET技术在神经元培养系统中定量检测新生蛋白合成速率,验证翻译水平对突触活动强度的实际响应。
Ribo-seq的核心价值:
稳态蛋白质组学往往难以捕捉神经元在突触活动期间瞬间发生的局部翻译变化。Ribo-seq在此解决了活动依赖性翻译的即时定量问题,清晰揭示了由于m6A降解路径受阻而堆积的转录本是如何在神经脉冲信号下迅速转变为功能性突触蛋白的。
核心发现:缺失YTHDF2显著抑制了突触功能相关mRNA的周转降解。这些未能及时清理的转录本在活动刺激下转化为极其活跃的翻译模板,产生更多的SEMA4B等关键蛋白,强化了突触传递效率,从而提升了小鼠的学习记忆能力。
该成果确立了YTHDF2介导的mRNA衰减是认知功能的负向调节阀,为改善记忆障碍提供了翻译调控水平的新靶点。
心血管医学

研究背景:
心脏发育伴随剧烈的能量代谢模式转换。NAT10同时具备RNA结合与乙酰转移活性,但既往研究对其驱动心脏成熟的具体功能域存在争议。该研究旨在理清究竟是哪种活性在维持心脏代谢基因表达和收缩力中占据主导地位。
研究思路与方法:
模型系统:研究通过心脏特异性Nat10敲除小鼠观察其对胚胎及产后发育的影响,并制备NAT10缺陷的人诱导多能干细胞衍生心肌细胞。通过对比体内超声心动图指标与体外细胞收缩功能,系统评估NAT10在不同物种系统中的功能保守性。
组学分析:对小鼠心脏组织进行了核糖体印迹测序与转录组的平行整合分析,利用delta TE计算方法筛选受调控基因。同步开展RIP-seq实验,在全基因组水平鉴定NAT10直接结合并保障丰度的候选靶标。
变异验证:构建了乙酰酶失活突变体G641E和RNA结合缺陷突变体K290A。通过在缺失模型中实施回补实验,判定NAT10发挥功能所依赖的具体结构域,以此解析心脏保护功能的分子基础。
Ribo-seq的核心价值:
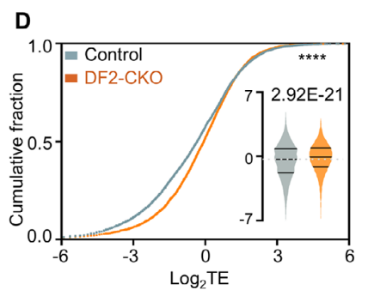
该研究中NAT10的缺失导致了大规模的蛋白丰度下降。通过Ribo-seq与RNA-seq的联合delta TE分析,研究者得以有效地将由mRNA降解导致的蛋白减少与由于翻译效率下降导致的蛋白减少区分开来,最终证明NAT10的保护效应主要发生在转录本稳定性层面。
核心发现:
NAT10主要通过RNA结合活性稳定脂肪酸氧化基因Acadm及收缩相关基因Ryr2的转录本。虽然它不显著改变全局翻译效率,但NAT10缺失引发的mRNA稳定性大幅下降共同导致了扩张型心肌病及产后死亡。
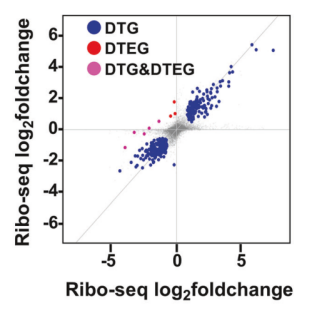 注:该图展示了绝大多数基因都在对角线上,证明全局翻译效率(TE)几乎没有改变
注:该图展示了绝大多数基因都在对角线上,证明全局翻译效率(TE)几乎没有改变
研究界定了NAT10作为RNA结合蛋白在维持心脏代谢平衡中的核心地位,为理解心脏发育障碍提供了翻译组学视角的解释。
肿瘤代谢

研究背景:
肝癌细胞必须重塑代谢与翻译模式以支持其快速增殖需求。该研究旨在寻找肝癌在翻译层面对特定氨基酸的极端依赖性,并基于此发现潜在的治疗漏洞。
研究思路与方法:
代谢示踪:运用同位素标记酪氨酸进行代谢流追踪实验,定量分析氨基酸在肿瘤细胞中进入分解代谢与蛋白合成途径的通量差异。通过对比不同模型验证肿瘤细胞如何通过上调tRNA相关因子来重塑氨基酸的利用路径。
翻译组监测:实施高深度的核糖体印迹测序以精确定位核糖体在mRNA上的实时位置及停留时间。通过分析足迹累积密度,捕捉酪氨酸限制条件下特定基因在密码子水平发生的核糖体行为异常,精确定位受翻译速度限制影响的关键代谢蛋白。
功能筛选:开展全基因组水平的CRISPR-Cas9协同致死筛选,在酪氨酸受限环境下鉴定影响癌细胞存活的基因。研究进一步结合药理学手段筛选与酪氨酸限制策略具有协同效应的临床药物,重点评估其在铁死亡通路中的治疗潜力。
Ribo-seq的核心价值:
常规质谱技术只能显示蛋白质的总量变化,却无法解释为什么氨基酸匮乏会导致特定的线粒体复合物受阻。Ribo-seq在此发挥了决定性作用,它捕捉到了TAC密码子处发生的显著核糖体停滞现象。为氨基酸匮乏直接限制翻译延伸速率提供了直接的物理证据,是其他组学技术无法实现的细节监测。
核心发现:
肝癌细胞通过MYC驱动上调合成酶YARS1和tRNA-TyrGUA,建立起一种极端依赖酪氨酸翻译的生存模式。研究提出的限制酪氨酸翻译可用性策略RTTA,会导致核糖体在TAC密码子处发生严重停滞,阻碍了线粒体复合物I及脂质调节物SCD1的翻译产出,诱发了氧化磷酸化衰竭与铁死亡。
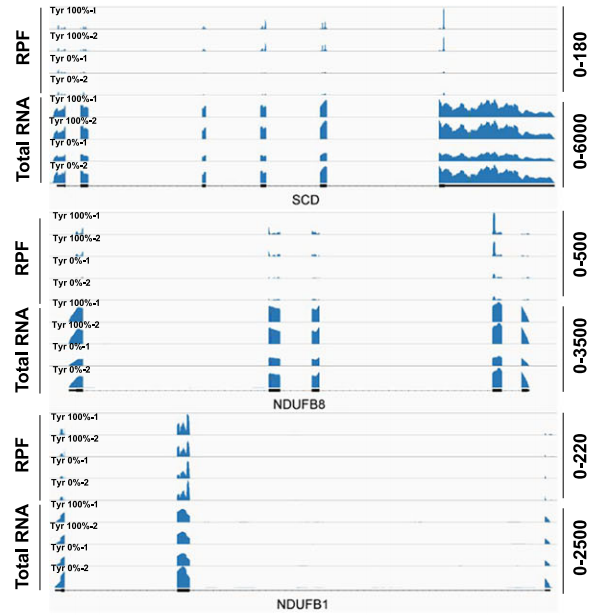
注:通过RPF Track清晰展示了在酪氨酸受限时,核糖体被固定在SCD1等基因上的特定位置
研究揭示了肝癌存在的酪氨酸翻译依赖性代谢漏洞,提出了通过精准干预特定密码子翻译速率来诱导癌细胞铁死亡的治疗新策略。
小结
这四篇文章只是一个截面。翻译调控的研究版图远不止于此——免疫细胞激活时的翻译重编程、衰老过程中核糖体保真度的下降、局部翻译在神经突触中的实时动态,都是正在被打开的方向。技术本身也在迭代,包括单细胞分辨率、三代全长的Ribo-seq在逐渐成熟。多组学的联合应用、面向临床微量样本的低起始量方案,让翻译组研究的边界持续外扩。从机制发现到靶点确认,翻译这一层的信息正在变得越来越不可绕过。


