
一文讲透cfDNA多模态,液体活检的前沿方向与研究路径
所谓的细胞游离DNA(cell-free DNA,cfDNA)是主要由凋亡或坏死细胞释放到血液、尿液等体液中的细胞外游离短DNA片段,这个cfDNA携带了来源细胞的遗传与表观遗传信息。cfDNA作为液体活检领域的核心研究对象,已在肿瘤早筛,产前检测、微小残留病灶(MRD)检测等领域展现出巨大应用潜力,至今在液体活检研究领域依然活跃。
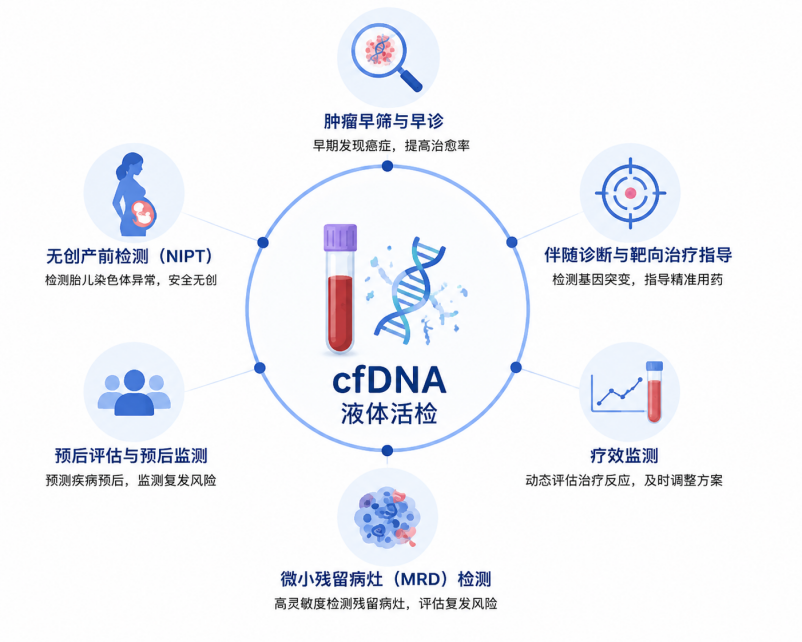
目前,cfDNA研究的主要生物标志物有基因突变、DNA甲基化和片段化模式,像大多数液体活检商品就是基于突变和甲基化。近十几年液体活检也经历了从最初的单一标志物到多维信息整合的范式转变,最常见的组合就是“突变+甲基化+蛋白”,其中AI驱动的深度学习整合cfDNA多模态特征(突变、甲基化、片段组学、组蛋白修饰、蛋白marker)的MCED显著提升了灵敏度与组织溯源准确率,这种策略也正在成为研究主流【1】。
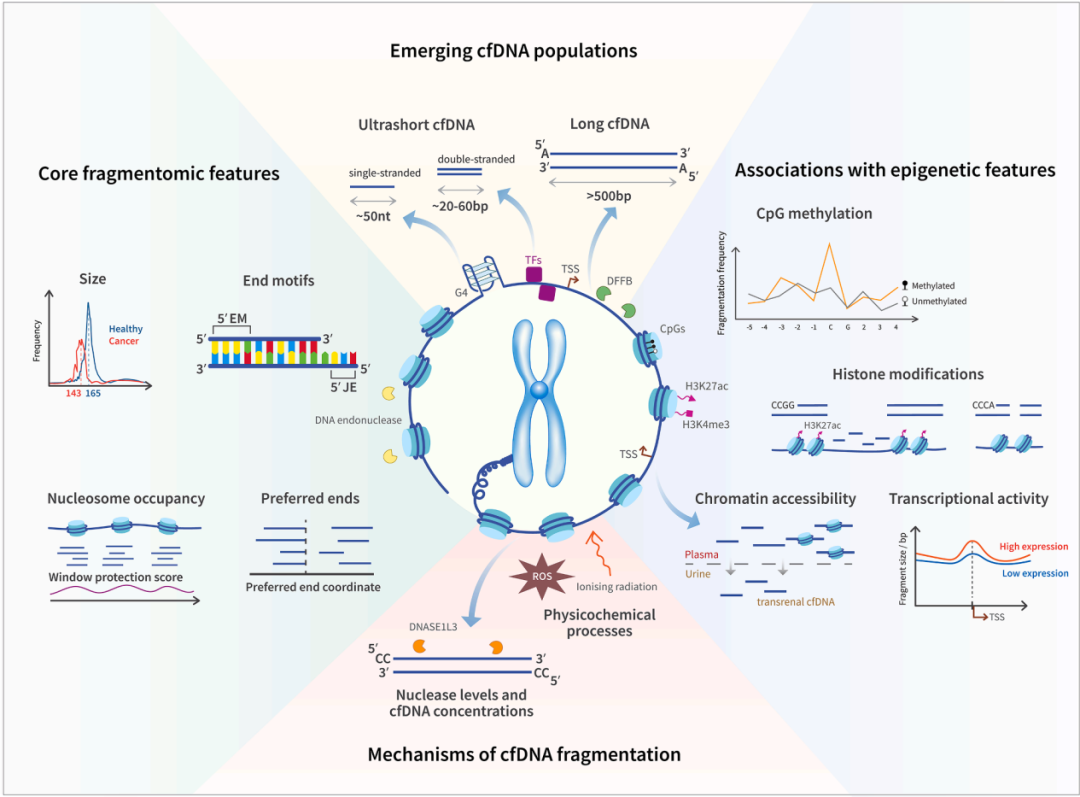
前面已提到当前cfDNA液体活检研究主要聚焦遗传变异(突变)、表观遗传(甲基化)、片段组学(片段特征)三个维度。这些年的深入研究也陆续发现这些维度蕴含关于细胞状态、组织来源和疾病进展的丰富信号。其中,cfDNA片段组学(cfDNA Fragmentomics)算是近年来液体活检领域发展最为迅速的前沿方向之一。其核心假设在于:不同组织来源的细胞在凋亡过程中,由于染色质结构和转录活性的差异,会产生具有特征性的cfDNA片段化模式。不过依赖单一维度在早筛、低肿瘤负荷、MRD和组织溯源中存在灵敏度不足、克隆性造血干扰、肿瘤异质性覆盖不足等问题,已不足以描绘肿瘤全貌。”表观遗传+片段组学+染色质状态”的多模态、多维度分析可能将会是cfDNA液体活检领域前沿热点方向之一。
👉cfDNA片段组学 ↔ DNA甲基化
-
DNA甲基化通过调控核酸酶活性影响cfDNA片段化。低甲基化增加核小体可及性,改变DNASE1L3等核酸酶的切割活性,导致cfDNA切割位点和大小分布发生变化;低甲基化区域产生更短的cfDNA片段【5】。
-
甲基化CpG位点在cfDNA片段末端富集。甲基化程度越高,片段在该位点断裂的偏好越强(最高2.4倍);肿瘤相关的低甲基化和基因高表达共同导致cfDNA片段变短【6】
-
片段组学特征可用于推断DNA甲基化状态。cfDNA片段组学特征(片段大小、末端偏好)可用于推断DNA甲基化状态,即从片段组学数据中"反推"甲基化组信息【7】;另外。差异甲基化位点周围的核小体定位模式与甲基化状态相关,HCC患者核小体模式振幅随肿瘤分期逐渐降低【8】。
-
多模态整合(甲基化+片段组学)显著提升癌症检测性能。SPOT-MAS方法同时分析cfDNA甲基化组和片段组,多模态整合显著优于单一维度,在多癌种早期检测中表现出更高的灵敏度和特异性【9】。
👉cfDNA片段组学 ↔ 基因突变
-
肿瘤来源ctDNA片段系统性偏短。携带肿瘤体细胞突变的ctDNA片段显著短于野生型cfDNA;通过富集短片段,EGFR T790M突变等位基因频率可提高约5倍,大幅提升早期癌症检测灵敏度【10】。携带突变的ctDNA片段比非突变cfDNA更短,体外尺寸选择富集短片段可显著提升ctDNA检测灵敏度【11】。
-
突变等位基因频率(VAF)与片段大小负相关。VAF越高,cfDNA片段越短,两者呈负相关,反映了肿瘤负荷与cfDNA片段化模式的内在联系【10-11】。
-
片段组学可区分肿瘤突变与克隆性造血(CH)突变。肿瘤来源cfDNA片段长度分布短于CH来源的cfDNA,片段大小分析可辅助区分两类突变来源,这对临床液体活检的假阳性控制具有重要意义【12】。
👉cfDNA片段组学 ↔ 组蛋白修饰
-
核小体保护决定cfDNA的基本片段特征 cfDNA的主峰约为166 bp,对应一个核小体核心颗粒(~147 bp)加上连接DNA的长度,这一现象的本质是组蛋白八聚体对DNA的物理保护【13】。通过深度测序血浆cfDNA,系统证明了cfDNA携带体内核小体足迹,核小体定位的细胞类型特异性可用于推断cfDNA的组织来源【14】。
-
活性组蛋白修饰(H3K27ac、H3K4me3)与cfDNA片段大小正相关。卢煜明团队发现循环核小体上H3K27ac和H3K4me3修饰水平与cfDNA片段大小存在正相关。活性修饰区域的cfDNA片段更长,而低修饰区域的片段更短,这一发现将组蛋白修饰状态与片段组学特征直接联系起来【15】。

-
cfDNA短片段保留转录因子结合足迹。短片段cfDNA(35-80 bp)在转录因子结合位点(CTCF、ETS、MAFK等)呈现保护性足迹,可推断体内TF结合状态;不同细胞类型的核小体定位差异可用于组织溯源【14】。Ulz等人开发了从cfDNA核小体占位模式推断504种转录因子可及性的方法,在前列腺癌中成功识别AR、NKX3-1、HOXB13等关键TF的活性变化,实现了癌症亚型分类【19】。
-
血浆cfDNA携带肿瘤特异性转录因子结合谱。血浆cfDNA中含有高分辨率的ER(雌激素受体)和FOXA1肿瘤结合谱,可识别乳腺癌ER驱动状态;TF-核小体动态模式反映肿瘤转录因子活性,可用于治疗监测【20】。
-
血浆cfDNA-蛋白复合物的蛋白质组学特征与癌症相关。血浆中循环cfDNA与蛋白质形成核小体-蛋白复合物(NPC),乳腺癌患者NPC的蛋白质组成与健康人显著不同,癌症患者NPC富集细胞骨架和蛋白复合物相关蛋白【21】。
-
单分子技术实现核小体修饰与蛋白标志物的同步检测。EPINUC技术在检测核小体组蛋白修饰的同时,可检测非分泌型肿瘤特异性蛋白(如突变型p53),实现cfDNA表观遗传与蛋白层面的多维整合,在不足1 mL血浆中即可完成多层次分析【22】。
👉DNA甲基化 ↔ 基因突变
-
甲基化与突变检测提供互补信息。在胰腺癌研究中发现,靶向DNA甲基化测序与突变检测并非总是相关,两者提供互补的肿瘤信息,联合使用可提升ctDNA检测灵敏度【23】。
👉组蛋白修饰 ↔ DNA甲基化
-
cfChIP-seq与cfMeDIP-seq提供互补的基因调控信息。H3K4me3 cfChIP-seq与cfMeDIP-seq从不同角度反映基因调控状态,两者联合描绘更全面的癌症表观遗传景观【16】。
-
伴随组蛋白修饰的核小体区域,其CpG甲基化状态共同决定cfDNA的表观遗传信息。组蛋白修饰区域的CpG甲基化状态影响cfDNA的片段化模式,两者协同编码了cfDNA的表观遗传信息,为从单一测序数据中同时推断甲基化组和组蛋白修饰状态提供了理论基础【7】。
👉cfDNA片段组学 ↔ 组蛋白修饰 ↔ DNA甲基化
这应该是目前研究较深入的三维关联研究,比如:
-
FRAGHA方法揭示片段组学-组蛋白修饰-甲基化的三维关联。他的核心机制:H3K27ac修饰区域具有更高的染色质可及性,而染色质可及性与DNA低甲基化状态相关;低甲基化区域的核小体排列更松散,核酸酶(尤其是DNASE1L3)切割偏好改变,最终导致该区域产生更短的cfDNA片段【1】。
-
甲基化-核酸酶-核小体-片段化四节点机制链。An等人在NC文章发现的机制:低甲基化→核小体可及性增加→DNASE1L3切割活性改变→cfDNA末端模式和片段大小改变【5】。
-
基因突变可通过改变全基因组甲基化模式影响cfDNA片段化。Noé等人发现IDH1突变导致全基因组高甲基化,进而使cfDNA片段末端CG富集增加、片段变长;反之,肿瘤相关低甲基化联合基因高表达共同导致cfDNA片段变短【6】。
👉片段组学 ↔ 组蛋白修饰 ↔ 基因表达
-
Snyder等人发表的研究成果表明,H3K36me3在基因体的cfChIP-seq信号与基因表达水平高度正相关【24】。Sadeh等人进一步将这一方法应用于肺癌,证明H3K36me3 cfChIP-seq可在血浆中直接推断肿瘤基因表达谱【24】。
-
cfDNA核小体占位模式可推断504种转录因子的结合可及性,且TF可及性评分与cfRNA基因表达水平显著正相关(p = 1.25×10⁻¹¹)【25】。
基于多维度特征的方法
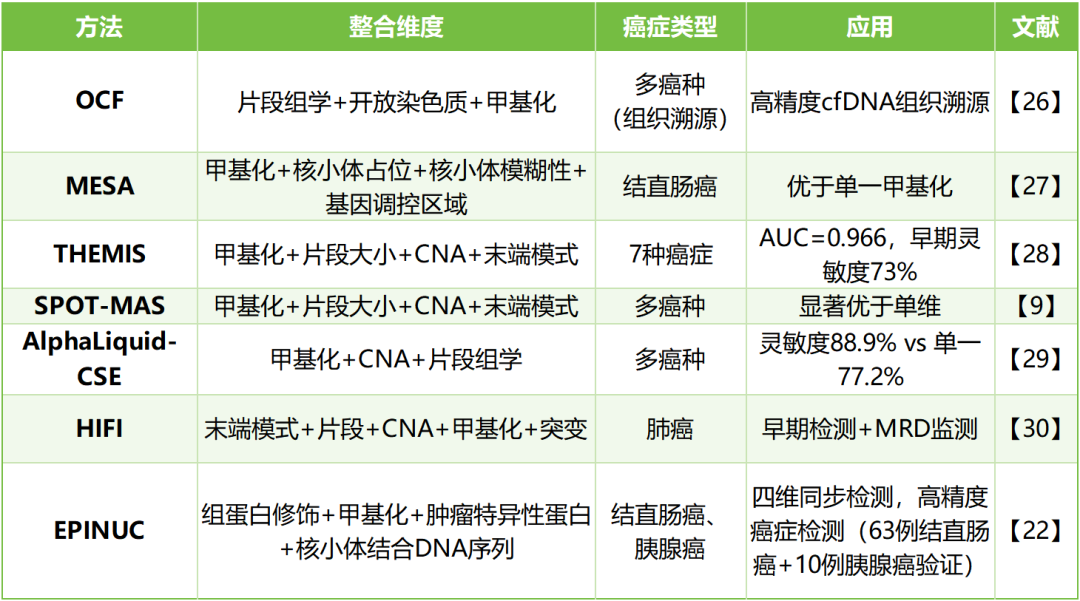
以上就是cfDNA液体活检研究领域中一些关键的多维度研究发现,还有其余更多的研究成果在这里就不一一追溯。关于cfDNA液体活检前沿研究动态可以关注星标我们公众号,表观生物会第一时间解读报道重大成果。
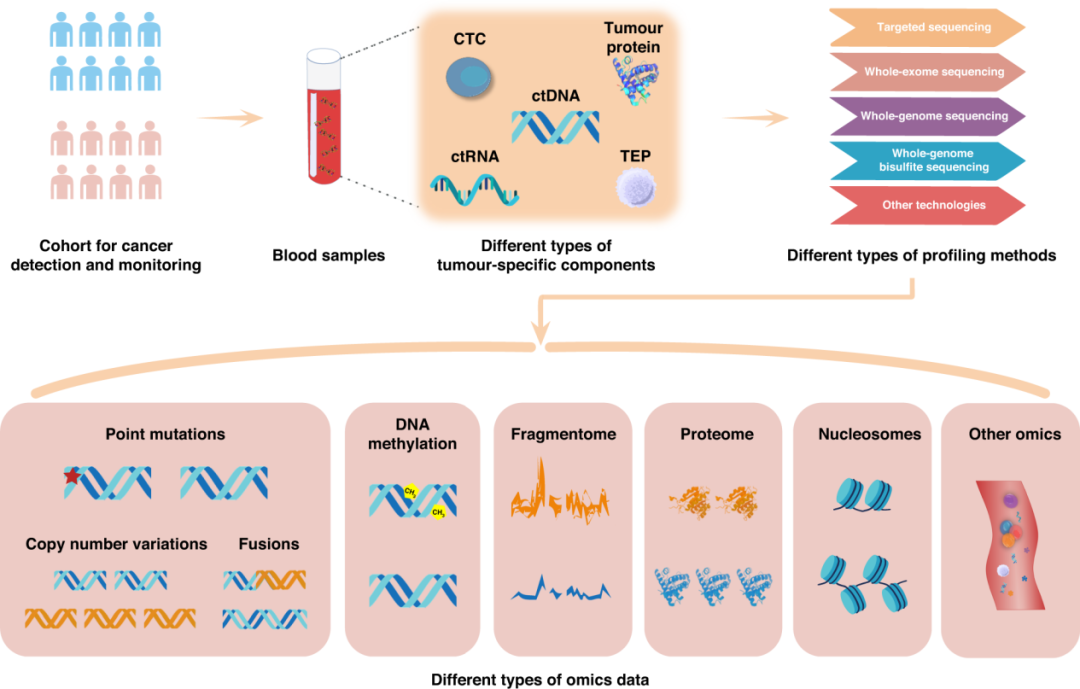
回到正题,那么既往的许多cfDNA液体活检研究,要实现多组学图谱的话,研究者往往需要开展多项组学实验,比如DNA甲基化需要通过WGBS、EM-seq等方法;片段组和基因突变需要通过WGS;cfDNA组蛋白修饰通过cfChIP-seq等。存在样本量需求高、分别建库、DNA转化损失等不足。
是否有更合适的、更全面的方案?这就来了:
表观生物最近刚推出了cfDNA片段组甲基化测序(FragMethyl-seq),FragMethyl-seq是我们基于illumina的5-base测序技术开发的cfDNA多模态分析技术方案:
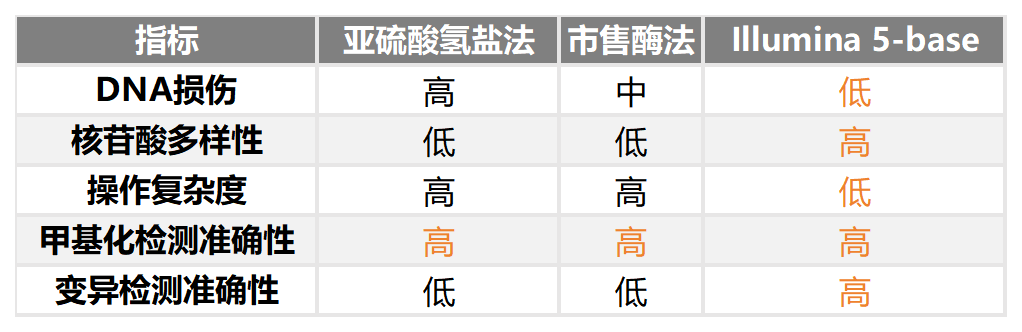
✔️一次建库、一次测序,同时获取DNA甲基化、片段化模式特征及基因突变等多维信息。无需拆分珍贵样本,避免批次差异。
✔️告别盐转法,采用一步酶促法,避免DNA降解损失,保留cfDNA片段完整与特征
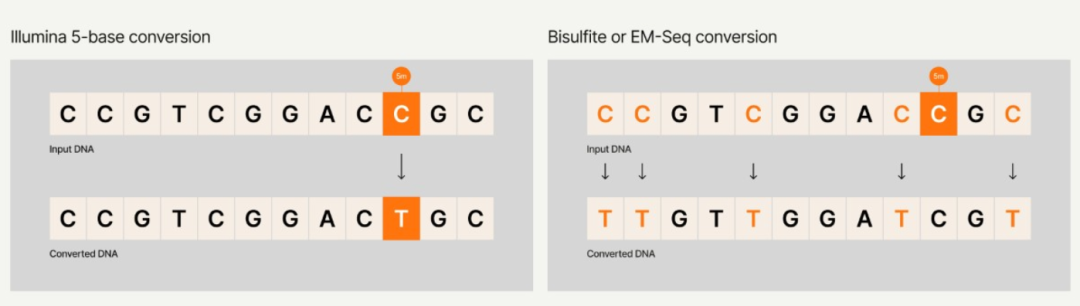
✔️极低起始量,低至1ng cfDNA即可启动,对珍贵样本友好,适合队列研究
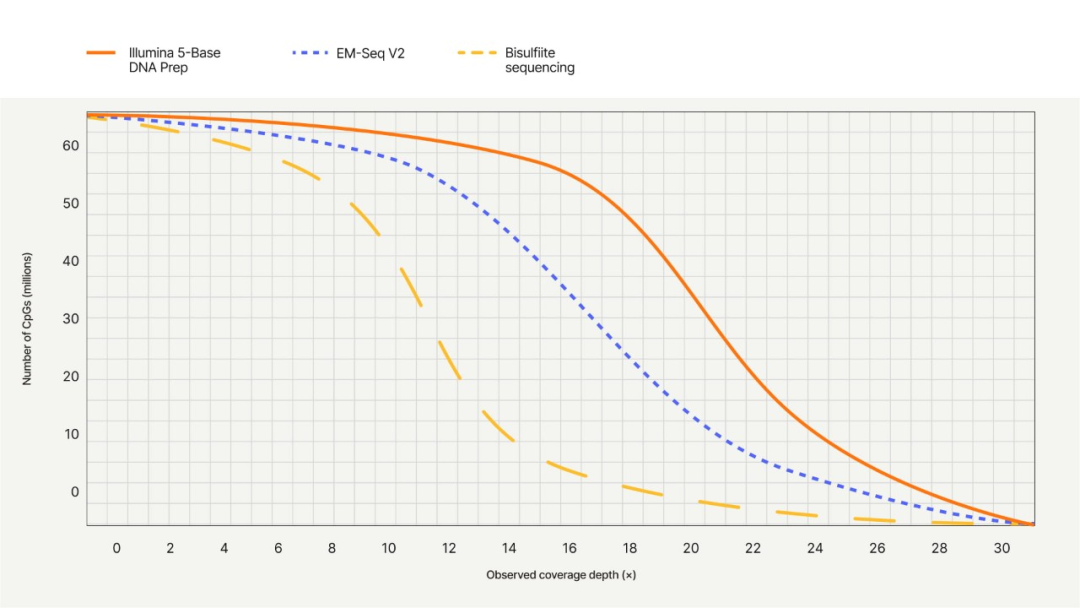
✔️单碱基分辨率,甲基化定位精确到单个CpG位点。Illumina 5-Base检测到的CpG位点中(900M 双端reads),10x以上覆盖深度的比例比EM-Seq v2多约20%,比亚硫酸氢盐测序多约68%。
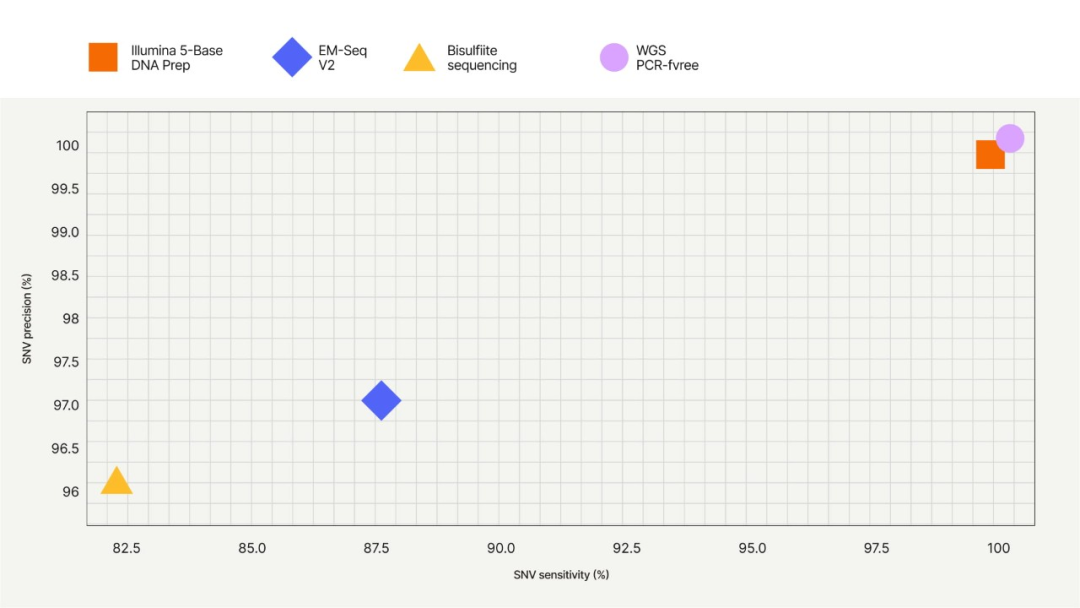
✔️检出SNV的灵敏度和准确性非常接近WGS(使用lllumina DNA PCR-Free Prep),高于EM-seq和亚硫酸氢盐测序。

去年,国家癌症中心发布《基于液体活检技术的多癌种联合筛查专家共识(2025版)》【3】。文章中专家共识意见明确指出甲基化联合片段组、cfRNA、蛋白质或代谢物具有提升检测性能的潜力。文章也列举一些灵敏度和特异性优异的成果案例,包括多模态产品,如DELFI(片段长度+核小体足迹)。共识还提及关于转化准确性、DNA损失的问题,强调高转化率、低偏差的建库体系。以甲基化为核心,片段组学等为补的多模态融合了,尤其针对MCED的多维AI建模,开展算法需求分析、建立泛化性评估体系与可解释性归因。FragMethyl-seq一次测序获取多维度信息正好是契合这些要素的
总之,FragMethyl-seq只为多癌种早筛、肿瘤组织溯源等液体活检场景的多模态分析提供了一套更全面、更高效的研究解决方案!
推荐上队列研究。特别是在多癌早筛、肿瘤组织溯源和分子分型研究中:
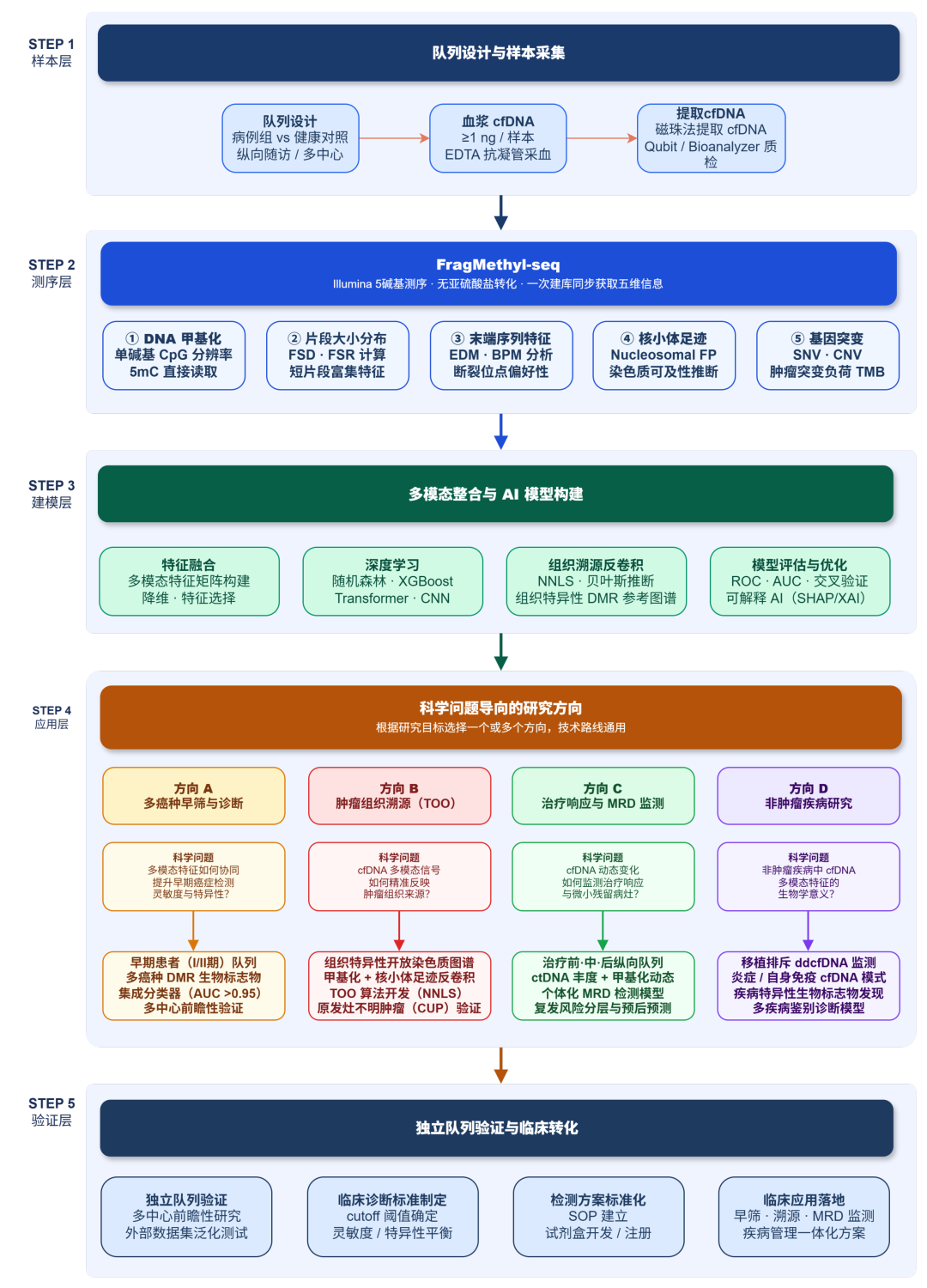
如果针对特定场景出发,可以参考以下几个基础的技术路径图:
1. 多癌种早筛与早期诊断
构建多维度特征矩阵,结合深度学习模型实现多癌种联合筛查。适合大规模队列早筛研究。
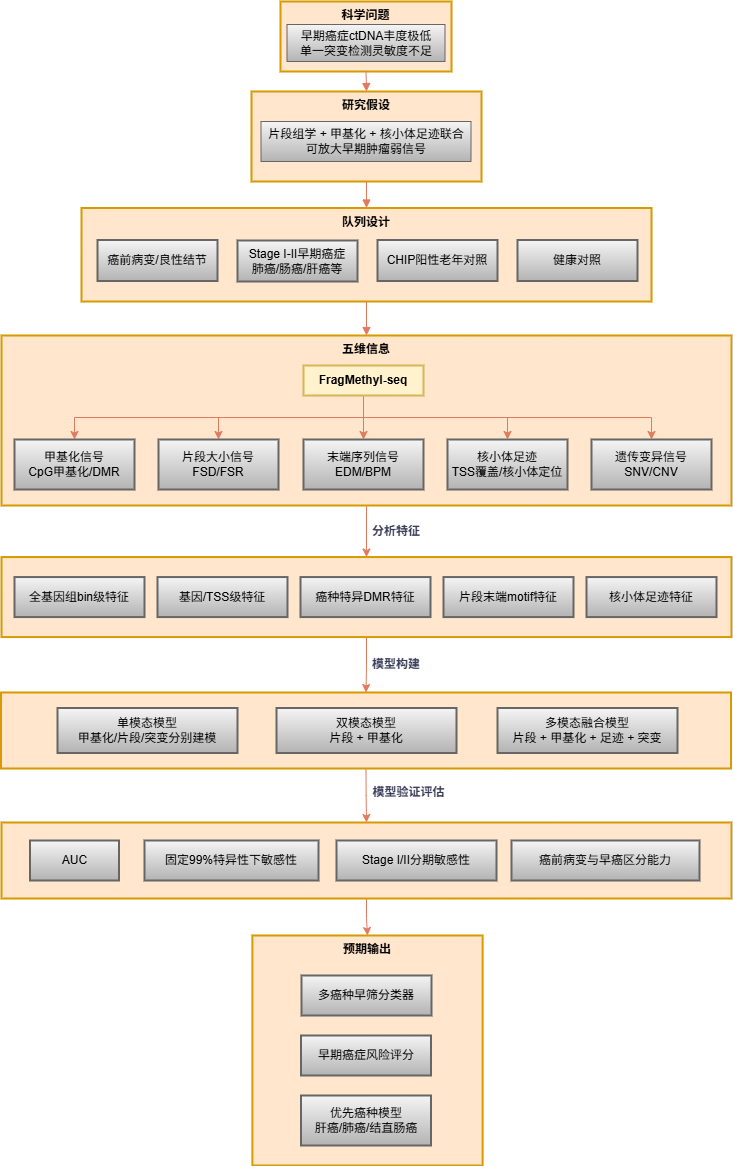
2. 肿瘤组织溯源与来源反卷积
当液体活检检出阳性信号时,明确肿瘤来源组织是指导后续临床检查的关键。甲基化反卷积分析可基于组织特异性DMR图谱,量化各组织对血浆cfDNA的贡献比例;结合片段组学的核小体足迹信号,进一步提升近邻组织的溯源分辨率。
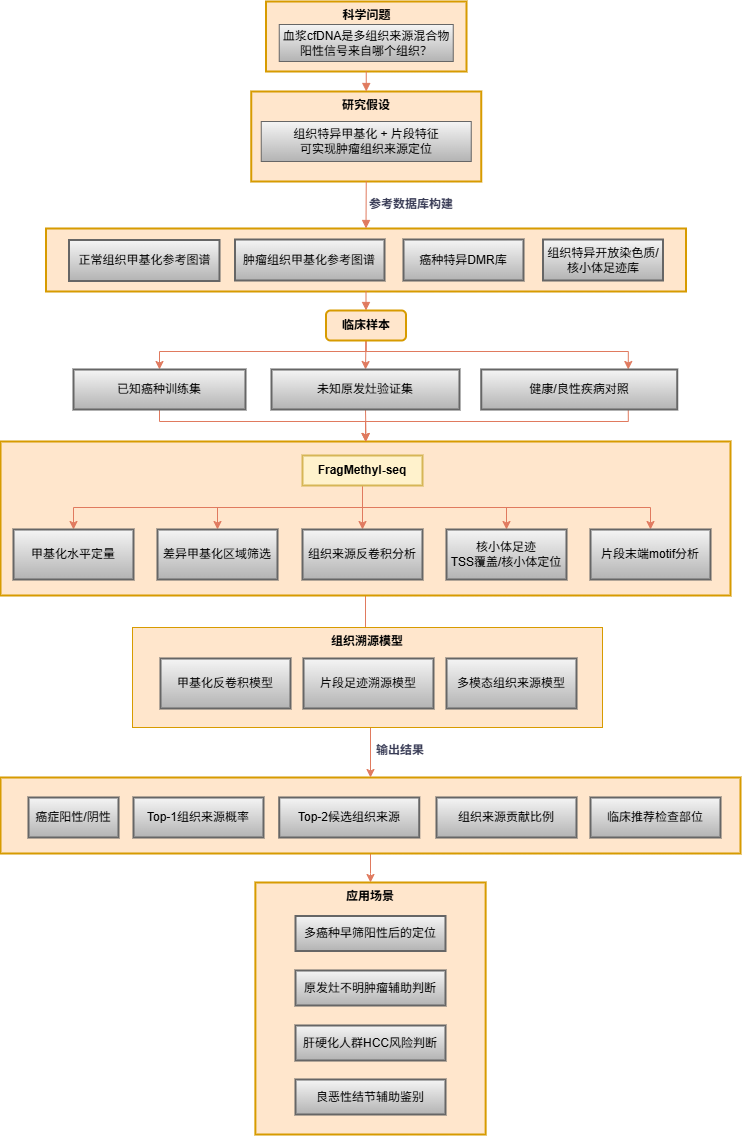
3. MRD监测与复发预警
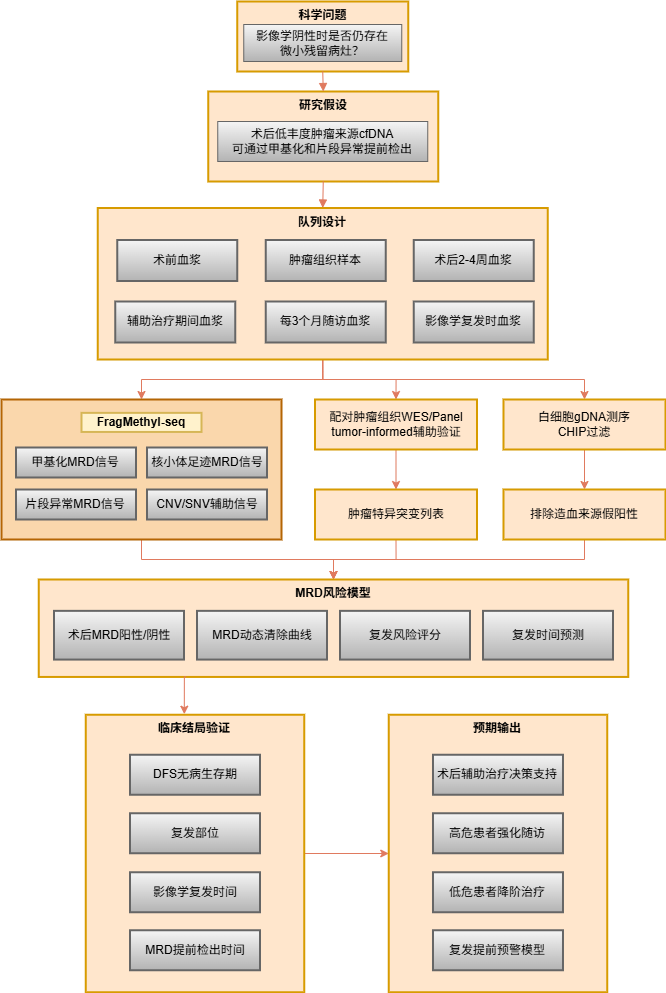
FragMethyl-seq低至1 ng起始量的建库能力,对术后随访的珍贵样本尤为友好。支持每2–3个月一次的密集动态随访采样。通过多时间点的甲基化水平变化(dM/dt)、肿瘤分数轨迹(dTF/dt)及片段组学特征联合分析,可构建早于影像学的复发预警模型。建议配合配对肿瘤组织panel及白细胞gDNA同步检测,以有效过滤CH引入的假阳性信号。
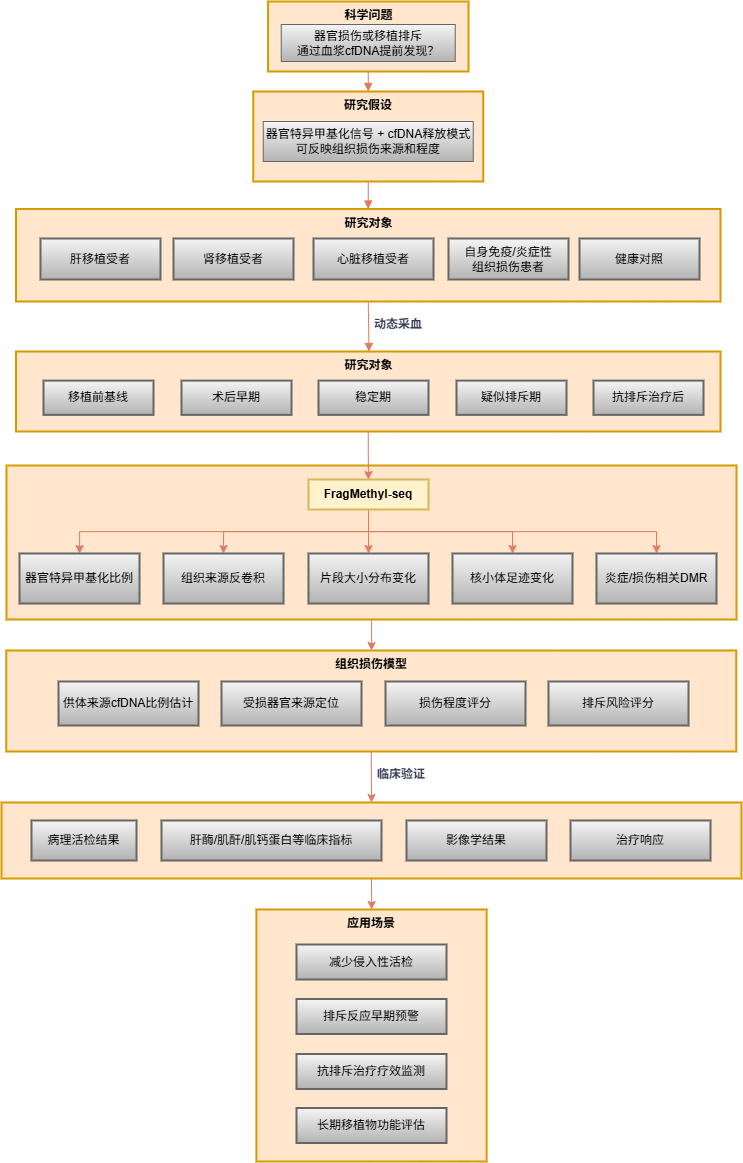
4. 移植排斥与组织损伤无创监测
器官移植后,受损或排斥的供体器官会向血浆中释放更多该器官来源的cfDNA。通过器官特异性甲基化反卷积,可定量血浆中供体来源cfDNA的比例变化;片段大小分布的异常偏移亦可作为组织损伤的辅助信号。
以上信息仅供cfDNA液体活检的多模态、多维度研究意向老师同学参考,内容请自行科学甄别。
此外,近期卢煜明院士在《Med》发表一篇文章“Cell-free DNA in 2030”【31】,回顾过去五年cfDNA领域的重大进展,并展望了2030年前的发展方向。其中有几点值得关注:
-
cfDNA生物学新发现:迄今最全面的细胞类型特异性甲基化图谱使组织溯源精度大幅提升【32】;研究发现濒死细胞中仅不到5%的DNA进入循环,这一清除悖论为解释不同部位肿瘤检出难度差异提供了新视角【33】。
-
cfDNA清除机制趋于清晰:经肾被动滤过、核酸酶降解、细胞主动摄取三条主要通路被进一步厘清;经肾清除的cfDNA优先来源于开放染色质区域,血小板被发现可主动富集携带肿瘤突变的cfDNA,为尿液诊断和血小板液体活检打开新窗口【34】。
-
主动干预cfDNA清除成为可能:通过抗dsDNA抗体 + 脂质体纳米颗粒启动剂策略,可在动物模型中将ctDNA检出量提升约60倍,有望破解早期癌症ctDNA极低丰度的瓶颈【35】。
-
新型cfDNA形态进入研究视野:约50 nt的"超短cfDNA"【36】和数千至数万bp的"长cfDNA"【37】分别被识别,结合长读长测序与Transformer/大语言模型架构的应用,cfDNA高维特征建模将迎来新一轮变革。
-
临床验证迈向硬终点:在约2万名无症状人群队列中,血浆EB病毒DNA的多模态片段组学分析可提前4年预测鼻咽癌发病风险,标志着cfDNA多模态分析正从"技术可行"走向"长期临床获益"【38】。
文章就此告一段落,这一期我们概述了cfDNA多维度的研究进展,列举cfDNA多模态研究技术路径。欢迎有意向的老师添加客服咨询。下一期cfRNA液体活检研究进展与思路方案,敬请关注。
参考文献
-
-
Bai J, Jiang P, Ji L, et al. Histone modifications of circulating nucleosomes are associated with changes in cell-free DNA fragmentation patterns. PNAS. 2024;121(42):e2404058121.
-
Tsui WHA, Jiang P, Lo YMD. Cell-free DNA fragmentomics in cancer. Cancer Cell. 2025;43(10):1792-1814.
-
陈万青, 陈可欣, 贺宇彤, 等. 基于液体活检技术的多癌种联合筛查专家共识(2025版)[J]. 中华肿瘤杂志, 2025, 47(7): 558-574.
-
Chen G, Zhang J, Fu Q, Taly V, Tan F. Integrative analysis of multi-omics data for liquid biopsy. Br J Cancer. 2023;128(4):505-518.
-
An Y, Zhao X, Zhang Z, et al. DNA methylation analysis explores the molecular basis of plasma cell-free DNA fragmentation. Nat Commun. 2023;14(1):287. Published 2023 Jan 18.
-
Noë M, Mathios D, Annapragada AV, et al. DNA methylation and gene expression as determinants of genome-wide cell-free DNA fragmentation. Nat Commun. 2024;15(1):6690. Published 2024 Aug 6.
-
Zhou Q, Kang G, Jiang P, et al. Epigenetic analysis of cell-free DNA by fragmentomic profiling. PNAS. 2022;119(44):e2209852119.
-
Zhu G, Jiang P, Li X, et al. Methylation-Associated Nucleosomal Patterns of Cell-Free DNA in Cancer Patients and Pregnant Women. Clin Chem. 2024;70(11):1355-1365.
-
Nguyen VTC, Nguyen TH, Doan NNT, et al. Multimodal analysis of methylomics and fragmentomics in plasma cell-free DNA for multi-cancer early detection and localization. Elife. 2023;12:RP89083. Published 2023 Oct 11.
-
Underhill HR, Kitzman JO, Hellwig S, et al. Fragment Length of Circulating Tumor DNA. PLoS Genet. 2016;12(7):e1006162. Published 2016 Jul 18.
-
Mouliere F, Chandrananda D, Piskorz AM, et al. Enhanced detection of circulating tumor DNA by fragment size analysis. Sci Transl Med. 2018;10(466):eaat4921.
-
Marass F, Stephens D, Ptashkin R, et al. Fragment Size Analysis May Distinguish Clonal Hematopoiesis from Tumor-Derived Mutations in Cell-Free DNA. Clin Chem. 2020;66(4):616-618.
-
Lo YMD, Han DSC, Jiang P, Chiu RWK. Epigenetics, fragmentomics, and topology of cell-free DNA in liquid biopsies. Science. 2021;372(6538):eaaw3616.
-
Snyder MW, Kircher M, Hill AJ, Daza RM, Shendure J. Cell-free DNA Comprises an In Vivo Nucleosome Footprint that Informs Its Tissues-Of-Origin. Cell. 2016;164(1-2):57-68.
-
Bai J, Jiang P, Ji L, et al. Histone modifications of circulating nucleosomes are associated with changes in cell-free DNA fragmentation patterns. Proc Natl Acad Sci U S A. 2024;121(42):e2404058121.
-
Baca SC, Seo JH, Davidsohn MP, et al. Liquid biopsy epigenomic profiling for cancer subtyping. Nat Med. 2023;29(11):2737-2741.
-
De Sarkar N, Patton RD, Doebley AL, et al. Nucleosome Patterns in Circulating Tumor DNA Reveal Transcriptional Regulation of Advanced Prostate Cancer Phenotypes. Cancer Discov. 2023;13(3):632-653.
-
Gezer U, Yörüker EE, Keskin M, Kulle CB, Dharuman Y, Holdenrieder S. Histone Methylation Marks on Circulating Nucleosomes as Novel Blood-Based Biomarker in Colorectal Cancer. Int J Mol Sci. 2015;16(12):29654-29662. Published 2015 Dec 11.
-
Rao S, Han AL, Zukowski A, et al. Transcription factor-nucleosome dynamics from plasma cfDNA identifies ER-driven states in breast cancer. Sci Adv. 2022;8(34):eabm4358.
-
Rao S, Han AL, Zukowski A, et al. Transcription factor-nucleosome dynamics from plasma cfDNA identifies ER-driven states in breast cancer. Sci Adv. 2022;8(34):eabm4358.
-
Tutanov O, Shefer A, Tsentalovich Y, Tamkovich S. Comparative Analysis of Molecular Functions and Biological Role of Proteins from Cell-Free DNA-Protein Complexes Circulating in Plasma of Healthy Females and Breast Cancer Patients. Int J Mol Sci. 2023;24(8):7279. Published 2023 Apr 14.
-
Fedyuk V, Erez N, Furth N, et al. Multiplexed, single-molecule, epigenetic analysis of plasma-isolated nucleosomes for cancer diagnostics. Nat Biotechnol. 2023;41(2):212-221.
-
Lapin M, Tjensvoll K, Edland KH, et al. Tumor-agnostic detection of circulating tumor DNA in patients with advanced pancreatic cancer using targeted DNA methylation sequencing and cell-free DNA fragmentomics. Mol Oncol. 2025;19(12):3535-3547.
-
Sadeh R, Sharkia I, Fialkoff G, et al. ChIP-seq of plasma cell-free nucleosomes identifies gene expression programs of the cells of origin. Nat Biotechnol. 2021;39(5):586-598.
-
Ulz P, Perakis S, Zhou Q, et al. Inference of transcription factor binding from cell-free DNA enables tumor subtype prediction and early detection. Nat Commun. 2019;10(1):4666. Published 2019 Oct 11.
-
Sun K, Jiang P, Cheng SH, et al. Orientation-aware plasma cell-free DNA fragmentation analysis in open chromatin regions informs tissue of origin. Genome Res. 2019;29(3):418-427.
-
Li Y, Xu J, Chen C, et al. Multimodal epigenetic sequencing analysis (MESA) of cell-free DNA for non-invasive colorectal cancer detection. Genome Med. 2024;16(1):9. Published 2024 Jan 16.
-
Bie F, Wang Z, Li Y, et al. Multimodal analysis of cell-free DNA whole-methylome sequencing for cancer detection and localization. Nat Commun. 2023;14(1):6042. Published 2023 Sep 27.
-
Kim SY, Jeong S, Lee W, et al. Cancer signature ensemble integrating cfDNA methylation, copy number, and fragmentation facilitates multi-cancer early detection. Exp Mol Med. 2023;55(11):2445-2460.
-
Li Y, Jiang G, Wu W, et al. Multi-omics integrated circulating cell-free DNA genomic signatures enhanced the diagnostic performance of early-stage lung cancer and postoperative minimal residual disease. EBioMedicine. 2023;91:104553.
-
Tsui WHA, Lo YMD. Cell-free DNA in 2030. Med. 2026;7(1):100963.
-
Loyfer, N., Magenheim, J., Peretz, A. et al. A DNA methylation atlas of normal human cell types. Nature 613, 355–364 (2023).
-
Sender R, Noor E, Milo R, Dor Y. What fraction of cellular DNA turnover becomes cfDNA?. Elife. 2024;12:RP89321. Published 2024 Feb 26.
-
Murphy L, Inchauspé J, Valenzano G, et al. Platelets sequester extracellular DNA, capturing tumor-derived and free fetal DNA. Science. 2025;389(6761):eadp3971.
-
Martin-Alonso C, Tabrizi S, Xiong K, et al. Priming agents transiently reduce the clearance of cell-free DNA to improve liquid biopsies. Science. 2024;383(6680):eadf2341.
-
Hudecova I, Smith CG, Hänsel-Hertsch R, et al. Characteristics, origin, and potential for cancer diagnostics of ultrashort plasma cell-free DNA. Genome Res. 2022;32(2):215-227.
-
Che H, Jiang P, Choy LYL, et al. Genomic origin, fragmentomics, and transcriptional properties of long cell-free DNA molecules in human plasma. Genome Res. 2024;34(2):189-200.
-
Lam WKJ, Kang G, Chan CML, et al. Fragmentomics profiling and quantification of plasma Epstein-Barr virus DNA enhance prediction of future nasopharyngeal carcinoma. Cancer Cell. 2025;43(4):728-739.e5.
>>
-

