
先驱转录因子IRF1如何让巨噬细胞越战越勇
巨噬细胞是先天免疫的核心执行者,而它的功能状态,很大程度上取决于染色质开合的动态调控。在静息状态下,巨噬细胞的基因组里其实潜伏着大片被核小体紧密包裹的区域。这些区域并非永久沉默,而是以潜伏增强子的形式蛰伏着,只等待特定的炎症信号来唤醒。一旦受到刺激,它们就能从零开始获得染色质可及性与调控活性,迅速驱动促炎基因程序的启动。
以往学界一直将谱系决定转录因子视为巨噬细胞染色质景观的主要架构,认为它们在髓系发育早期就预先铺好了增强子库,后续的信号依赖性转录因子只需“按图索骥”即可。但有一个关键问题:面对IFNγ这类强效促炎信号,是否存在一类不依赖既有地基、能够直接侵入封闭染色质的先驱转录因子?如果有,它又是如何完成这一过程的?
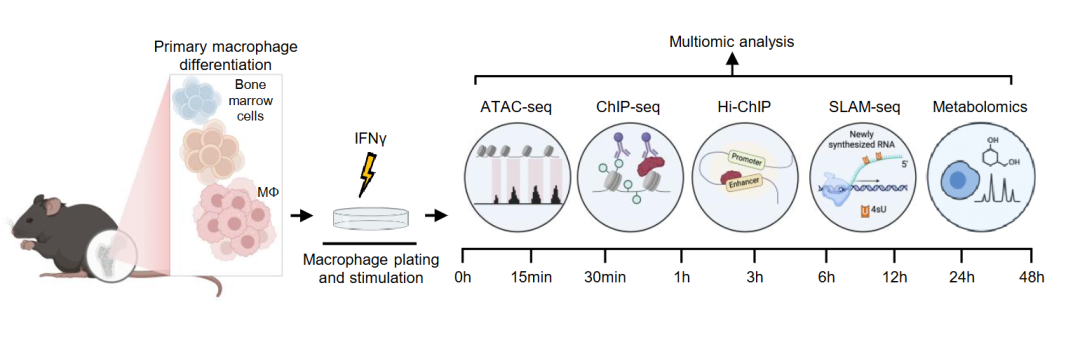
要拆解这个机制,需要通过哪些手段?是否有一套适配的多组学的交叉验证?刚好,最近看到一篇案例,麦吉尔大学David Langlais团队在BioRxiv上传的一篇预印本,提供了一个漂亮的整合分析范本。这项研究综合运用ATAC-seq、ChIP-seq、HiChIP及SLAM-seq等多组学技术,并系统证明IRF1是IFNγ驱动巨噬细胞激活过程中具有先驱活性的转录因子。IRF1能够直接侵入封闭染色质,驱动潜伏增强子从头建立,并通过长程染色质互作激活免疫效应、染色质调控和细胞代谢相关转录程序;同时,IRF1协调有氧糖酵解代谢切换,依赖BRG1-SWI/SNF染色质重塑复合体实现增强子开放,且所建立的增强子状态在IFNγ撤除后持续存在,形成可遗传的表观免疫记忆。这项工作的研究思路与方案设计值得作为相关研究领域的参考。

研究技术路线
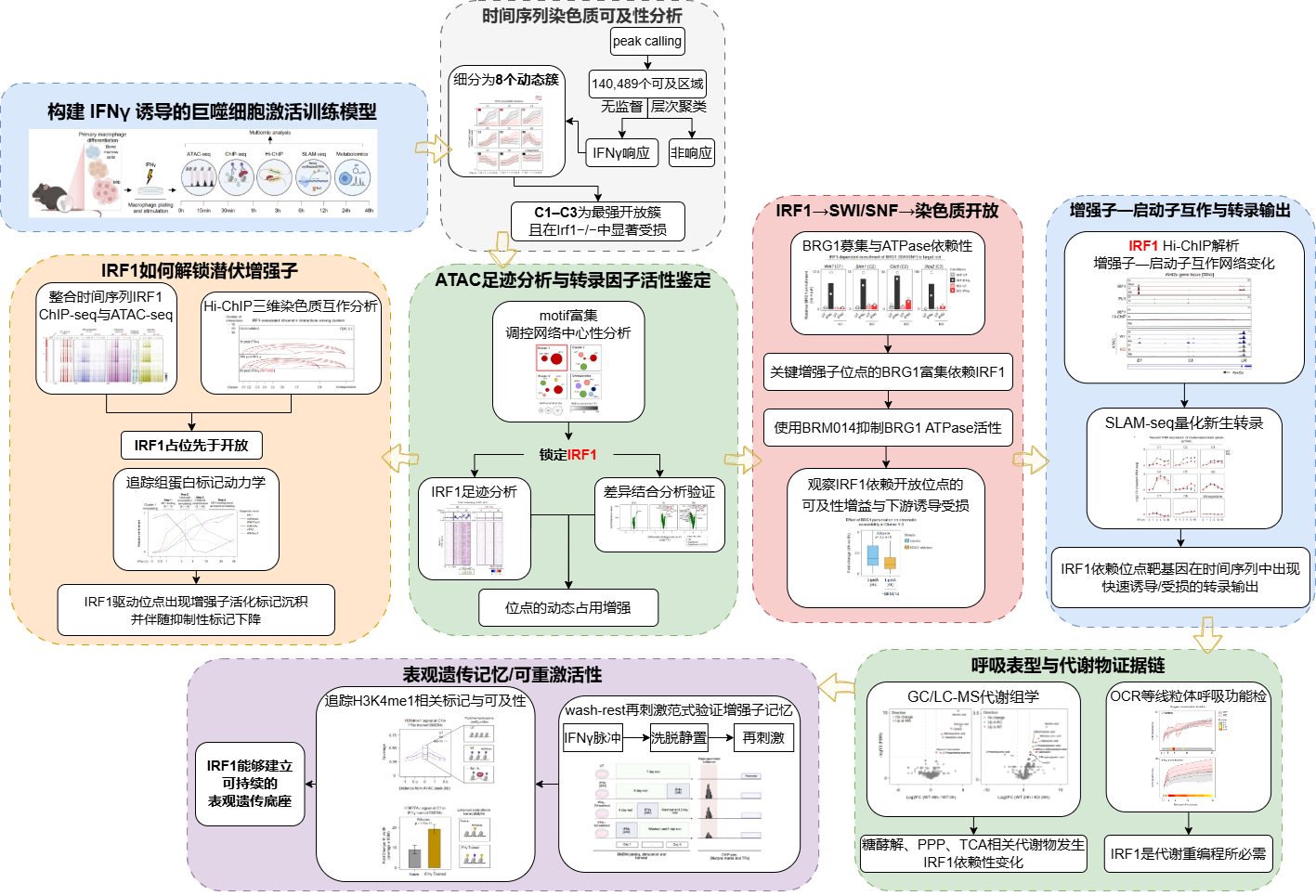
关键研究结果
1、IRF1独立于PU.1驱动IFNγ响应性染色质开放
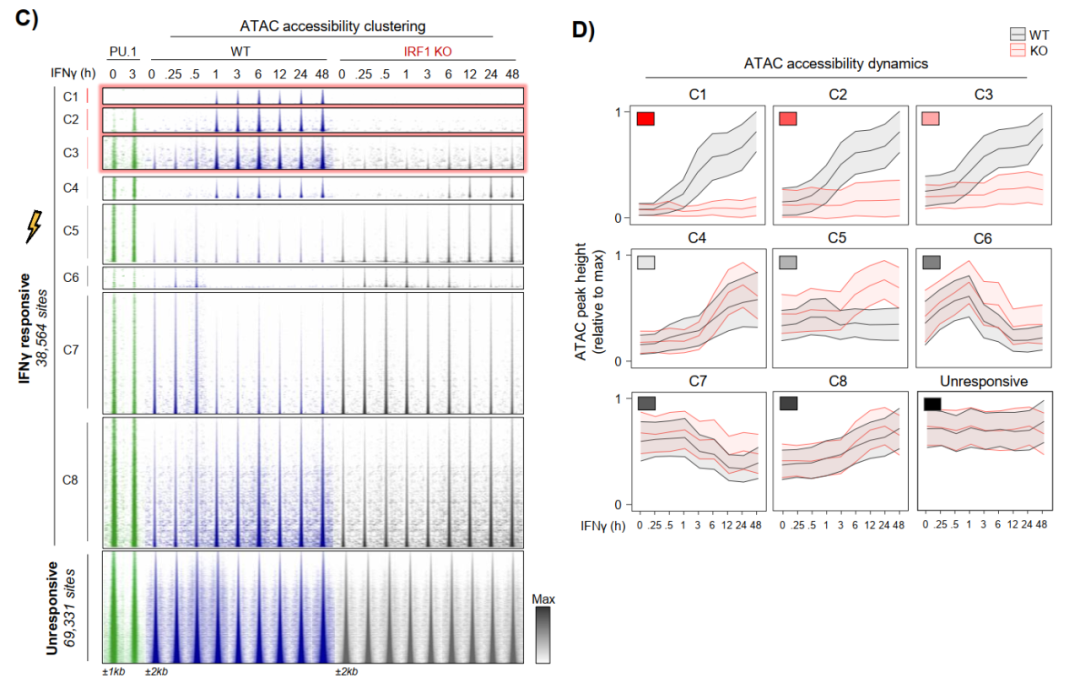
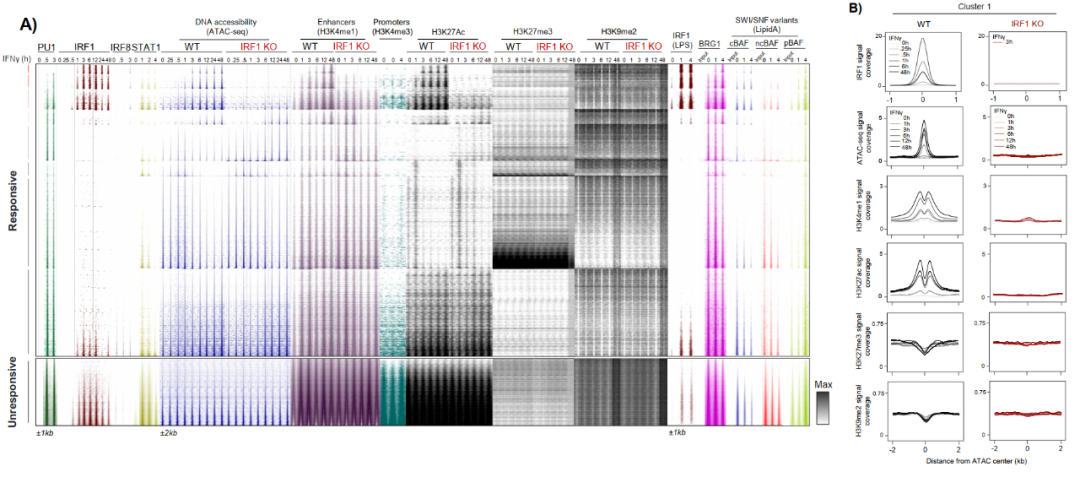
文章通过ATAC-seq鉴定出140,489个可及性区域,其中38,564个呈IFNγ响应性,经k-means聚类分为C1-C8共8个簇。C1、C2、C3簇在IFNγ刺激后表现出最强的染色质开放,且在IRF1 KO细胞中显著受损。
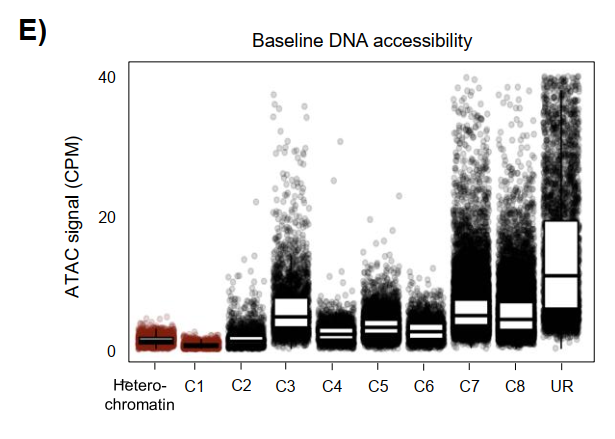
尤其是C1簇,在无IFNγ刺激时染色质可及性低至接近异染色质水平,且完全依赖IRF1获得开放性。而且C1位点缺乏经典先驱因子PU.1的占据信号,表明IRF1在这一区域的先驱活性完全独立于PU.1,拓展了巨噬细胞染色质可塑性的范畴。
2、IRF1具备经典先驱因子特征:结合封闭染色质并建立潜伏增强子
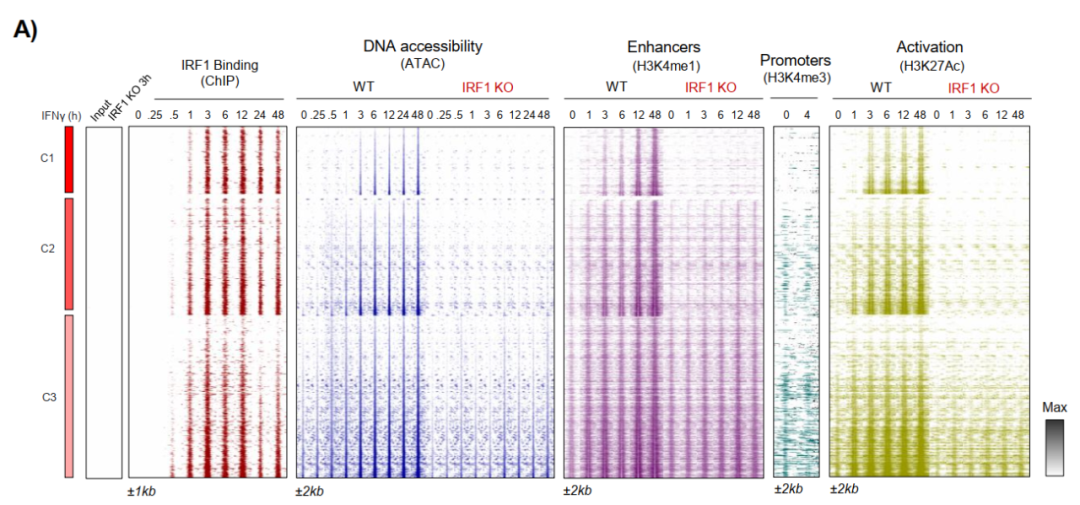
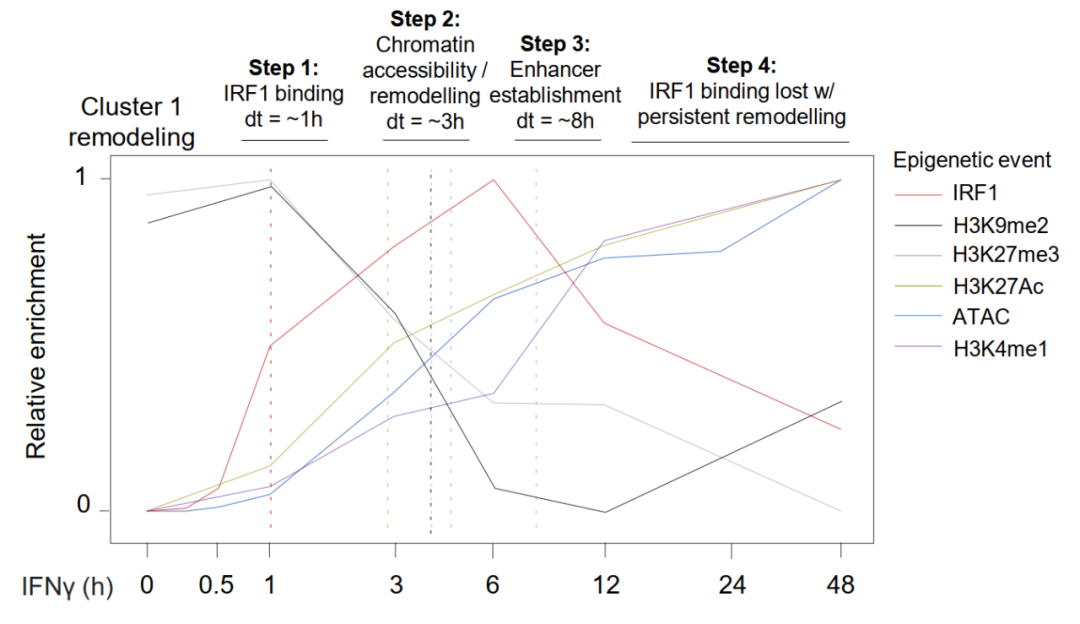
IRF1 ChIP-seq显示,IRF1结合早于ATAC-seq可检测到的染色质开放约2小时(C1位点IRF1结合半时约1小时,ATAC开放约3小时),这是先驱因子侵入封闭染色质的核心证据。
C1位点在基线状态缺乏H3K4me1和H3K27ac增强子标志,随IFNγ刺激逐步获得这些活性标志,同时丢失抑制性标志H3K9me2和H3K27me3,且上述变化完全依赖IRF1。
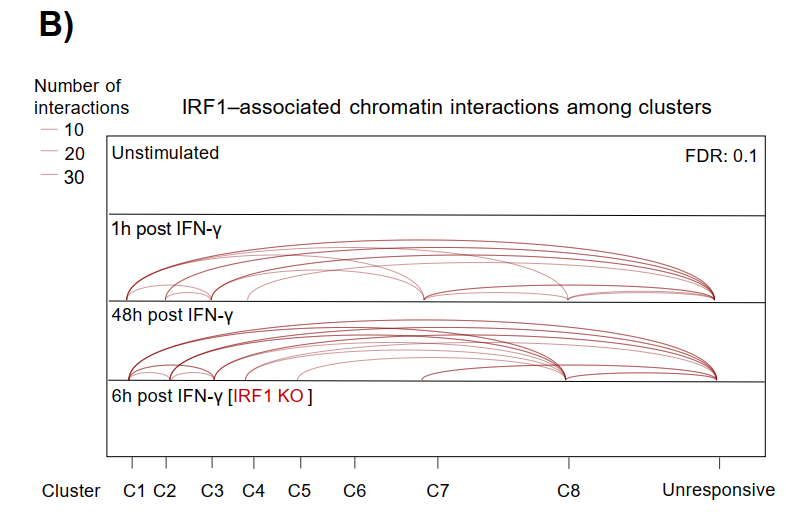
通过HiChIP测序进一步显示C1-C3簇增强子与基因启动子之间形成大量长程染色质互作,而这些互作在IRF1 KO细胞中消失,证明IRF1所建立的潜伏增强子在功能上整合入基因调控网络。
3、高密度IRF1 motif阵列决定先驱活性强度
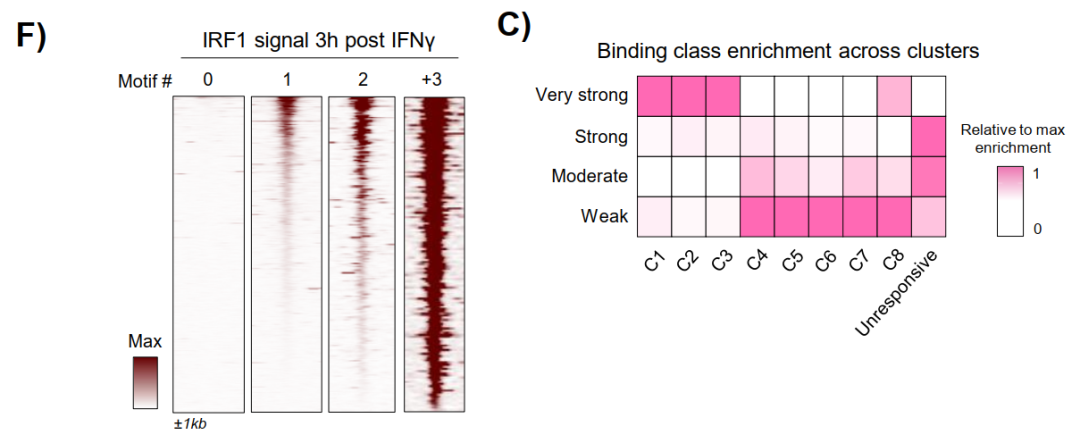
对55,416个IRF1结合位点的系统分析揭示,IRF1占据强度与其motif数目呈正相关,最强结合位点的C1-C3富集最为显著。
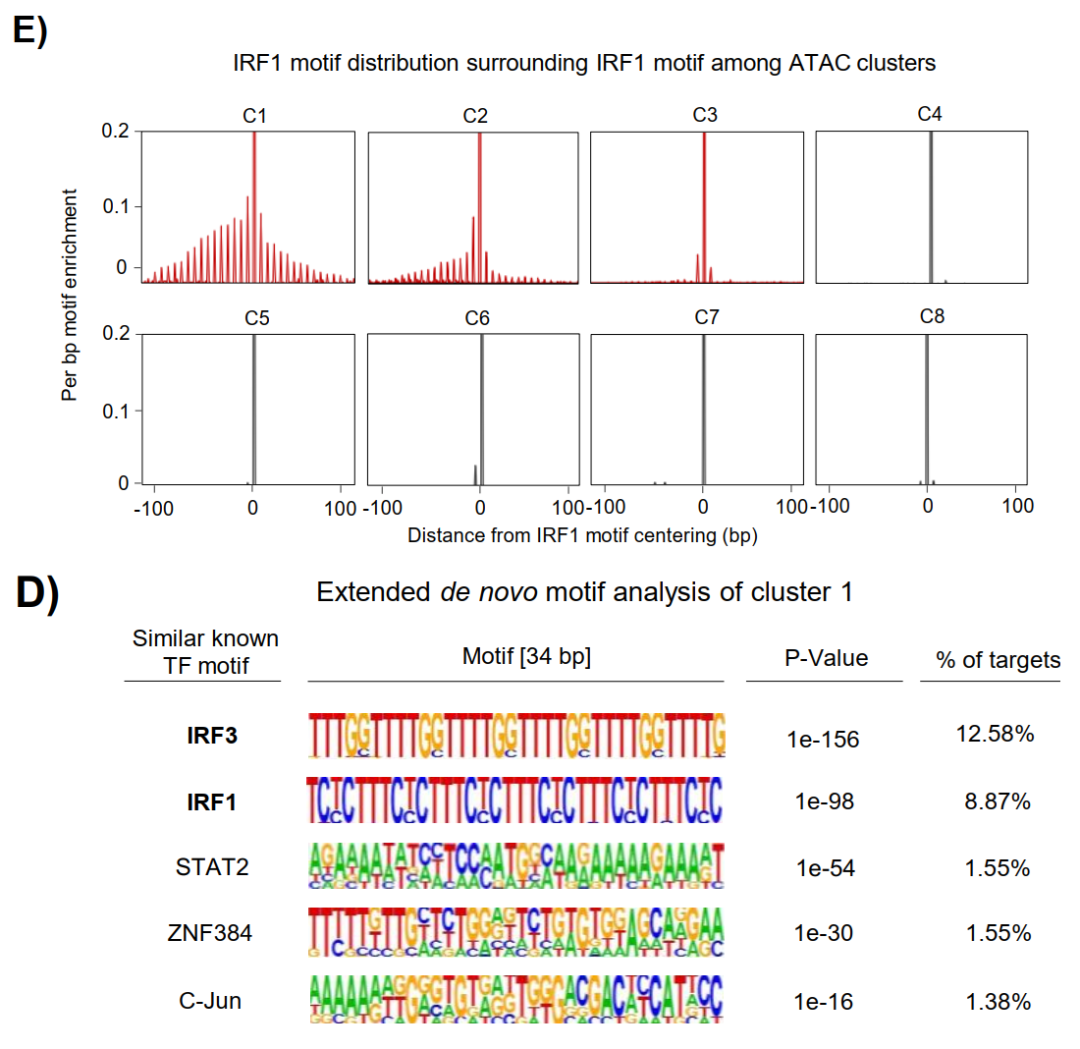
C1位点存在有序间隔排列的多重IRF1 motif阵列,且motif数目越多,HiChIP检测到的增强子-启动子成环频率越高,约至15个motif后趋于饱和。
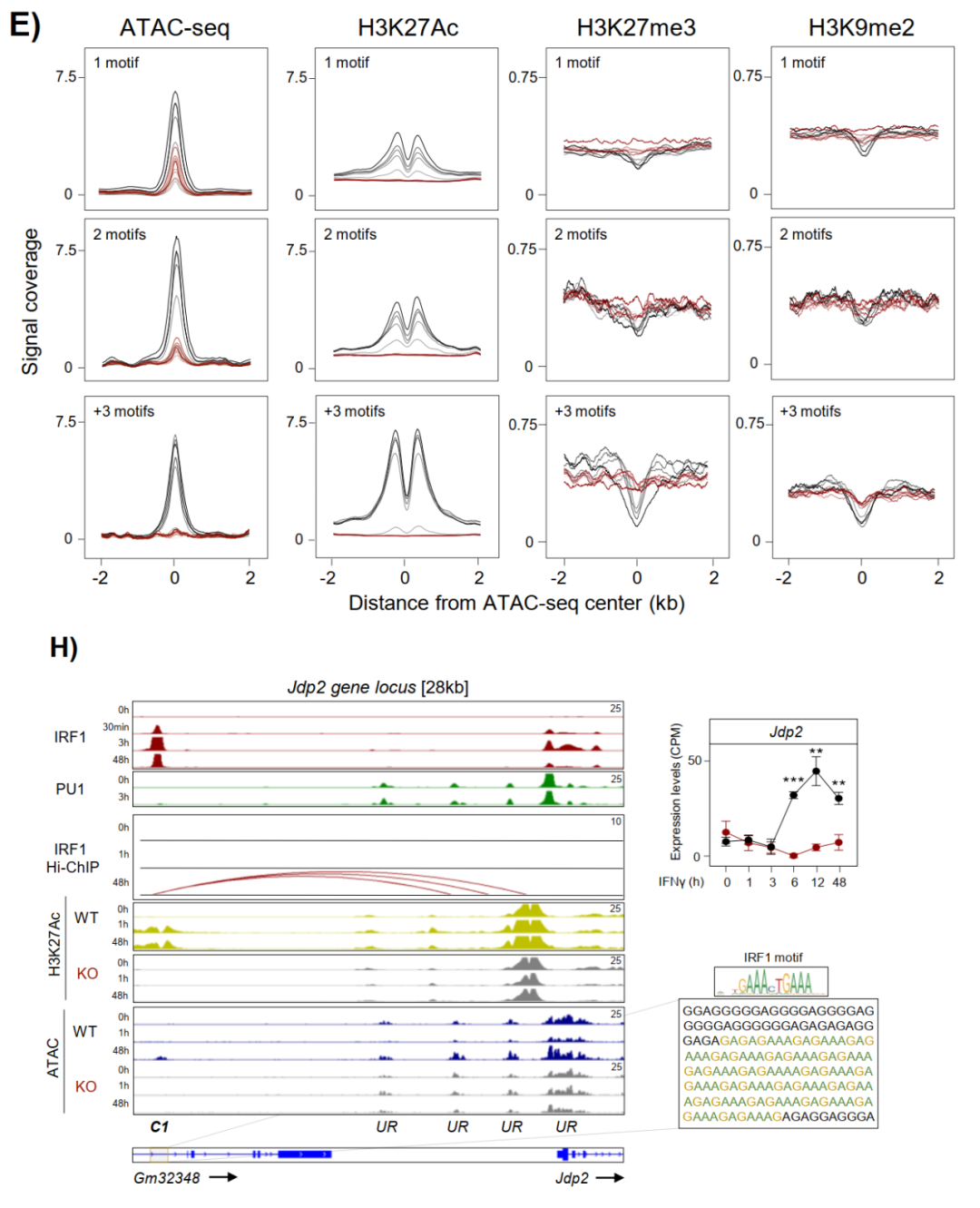
在IRF1 KO细胞中,多motif位点完全无法获得染色质开放或H3K27ac沉积,Jdp2位点的IGV图直观展示了motif阵列与IRF1先驱活性的对应关系。
4、IRF1-SWI/SNF轴驱动增强子重塑与靶基因转录
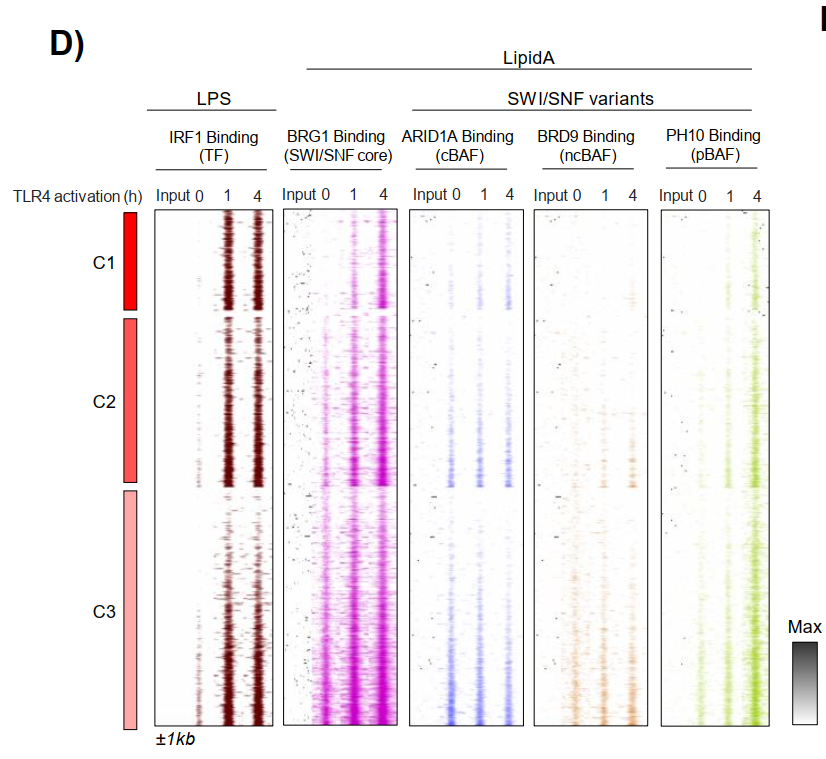
IRF1招募BRG1-SWI/SNF复合体是其先驱活性的关键分子机制。利用已发表LPS/Lipid A刺激数据集,研究显示SWI/SNF核心亚基BRG1、ARID1A(cBAF)和PHF10(pBAF)在IRF1结合后1小时内即被招募至C1-C3增强子。
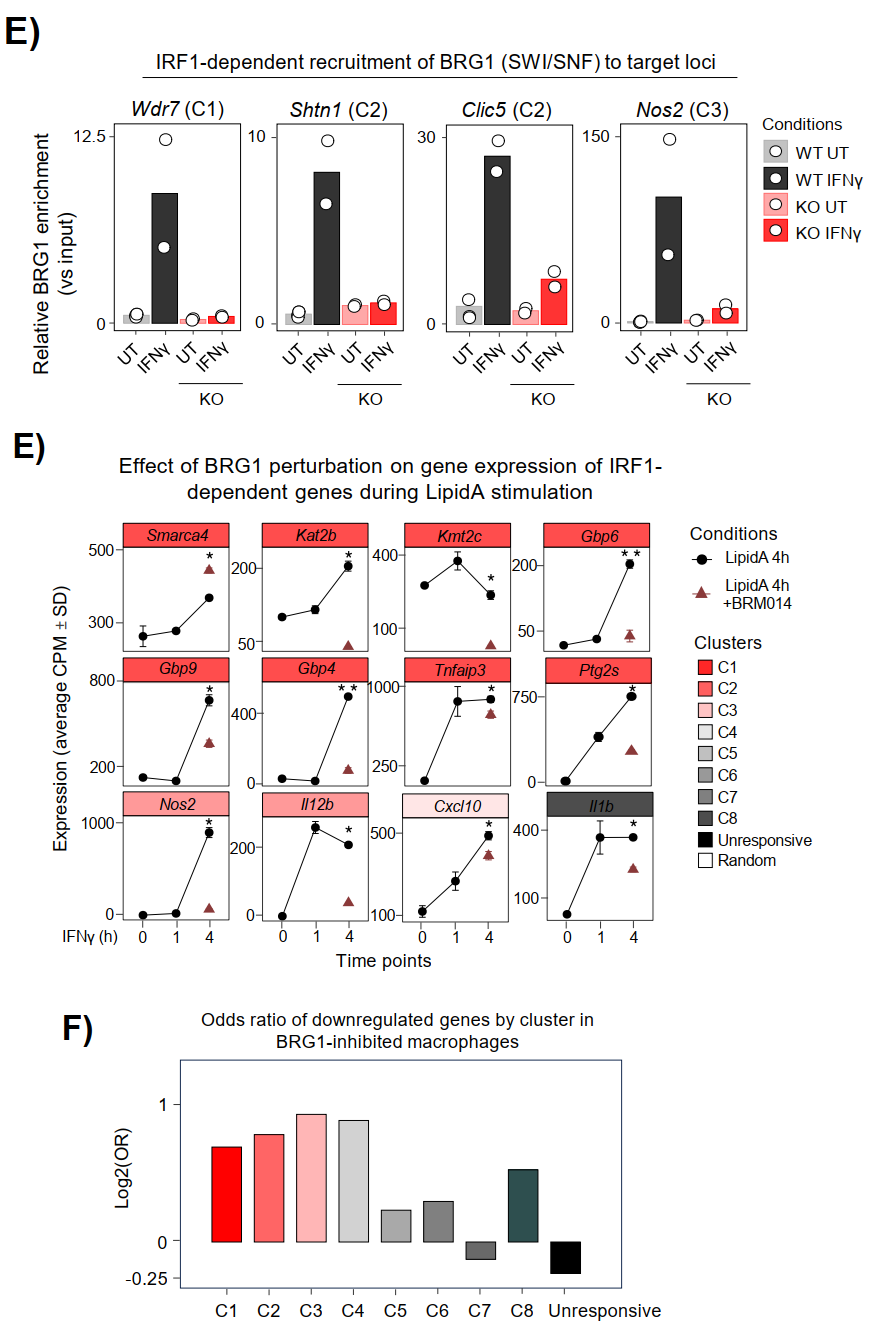
在IRF1 KO细胞中,IFNγ诱导的BRG1富集在Wdr7、Shtn1、Clic5、Nos2等位点完全消失,药理学抑制BRG1(BRM014)显著减弱Lipid A诱导的C1-C3染色质开放。转录层面,BRG1抑制选择性阻断IRF1依赖性靶基因包括Smarca4、Kmt2c、Gbp6的表达激活,且IRF1依赖簇受到不成比例的抑制。
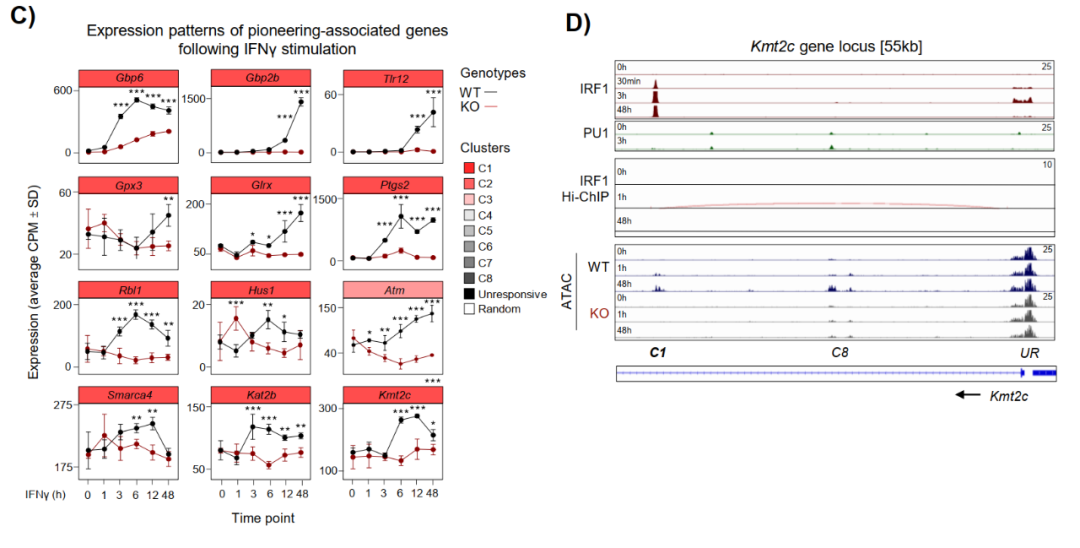
SLAM-seq结果还揭示IRF1通过建立潜伏增强子直接控制例如Smarca4、Kat2b、Kmt2c等多个表观遗传调控因子的转录,形成正反馈放大回路。
5、IRF1协调IFNγ驱动的代谢重编程
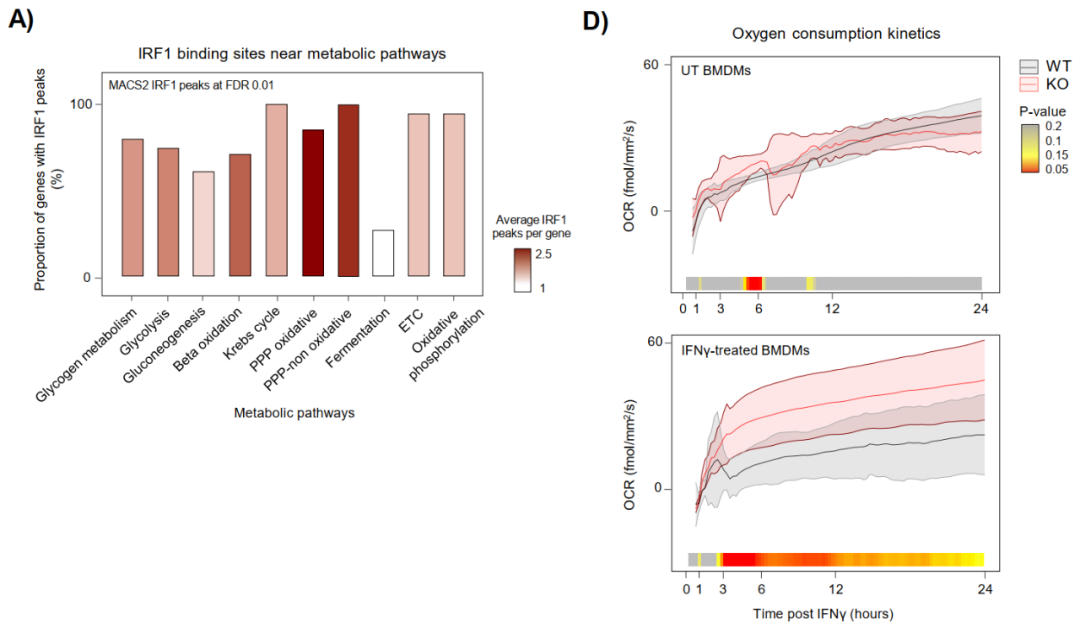
代谢组与转录组整合分析证明IRF1是IFNγ代谢切换的直接转录驱动者。IRF1结合广泛分布于糖酵解、PPP(磷酸戊糖途径)和TCA循环相关基因位点,IRF1 KO细胞在IFNγ刺激后无法实现从氧化磷酸化(OXPHOS)向有氧糖酵解的切换,持续维持较高OCR。
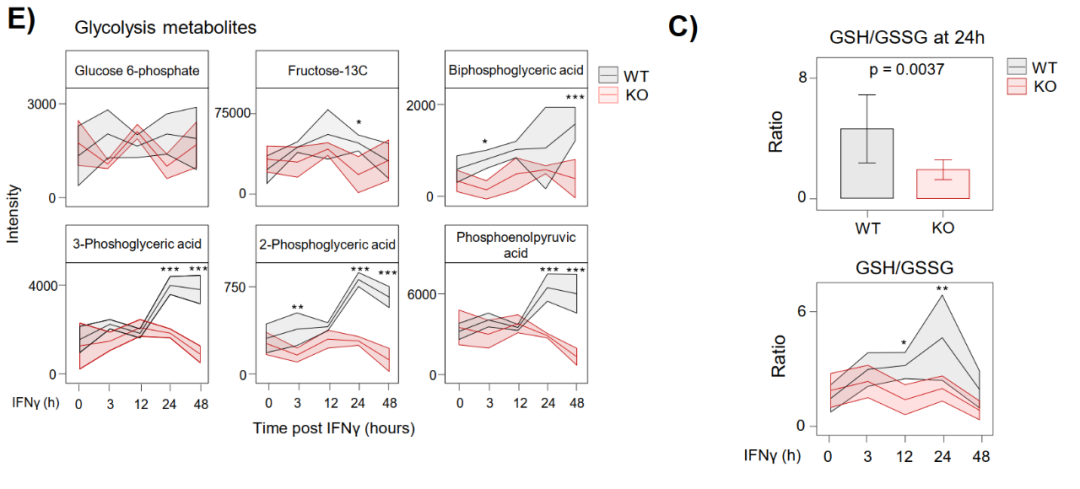
靶向代谢组学检测到WT细胞中3-磷酸甘油酸、2-磷酸甘油酸、2,3-二磷酸甘油酸和磷酸烯醇式丙酮酸等晚期糖酵解中间体显著积累,而KO细胞中缺失。TCA方面,IRF1在IFNγ刺激后15分钟内即结合Acod1启动子,驱动其强烈表达itaconate(及衣康酸)积累;谷胱甘肽氧化还原比值(GSH:GSSG)在WT细胞中于24小时时点显著高于KO,反映IRF1通过PPP增强NADPH生成和氧化还原缓冲。
6、IRF1建立持久性表观免疫记忆
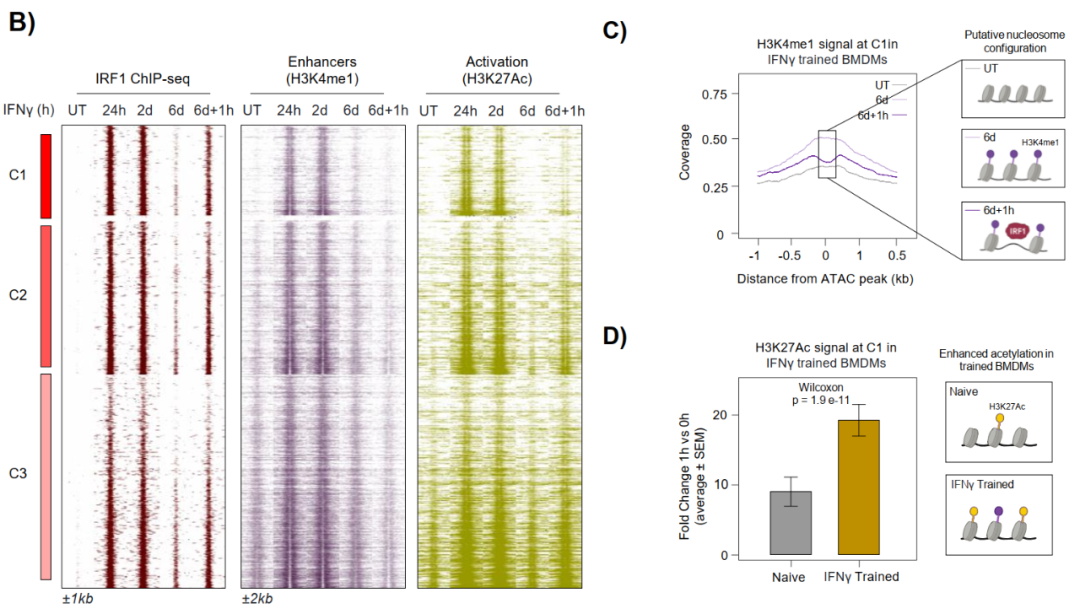
Washout实验显示,IFNγ撤除6天后,C1-C3位点H3K4me1标志持续保留,但活化标志H3K27ac回归基线。1小时再刺激后,这些"训练"增强子快速重获H3K27ac,响应速度显著快于初次刺激。
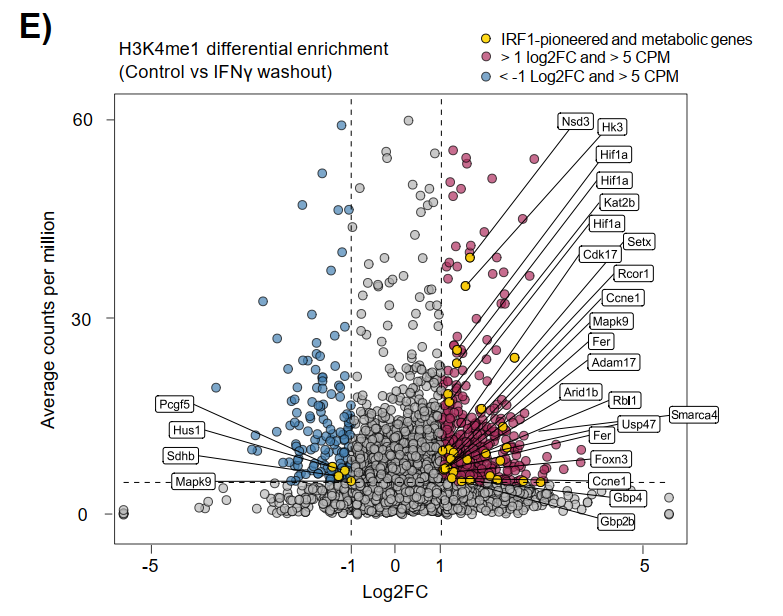
持续保留H3K4me1的位点毗邻代谢(Hk3、Hif1a)、染色质重塑(Kat2b)、细胞周期(Rbl1、Ccne1)和防御功能(Gbp2b、Gbp4)等相关基因,提示IRF1的先驱活性在初次炎症暴露后留下功能性表观遗传印记,使巨噬细胞具备更敏捷的再激活能力。
小结
在分子机制层面,作者将IRF1的时序性作用模式与FOXA1等经典先驱因子类比,提出高密度IRF1 motif阵列通过协同诱导DNA弯曲不稳定化核小体结构并促进SWI/SNF招募的假说,这一机制与OCT4/SOX2的协同motif效应具有功能相似性。IRF1驱动的TCA循环重塑不仅产生衣康酸等免疫代谢效应物,富马酸、乙酰-CoA等中间代谢物亦可作为染色质修饰酶的底物,形成代谢与表观遗传状态的正向强化回路,为持久性表观记忆提供代谢基础。在训练免疫框架内,BCG疫苗接种后IRF1上调及β-葡聚糖训练中性粒细胞中IRF1 motif高度富集等证据,均支持IFNγ-IRF1轴作为表观遗传训练核心通路的定位。未来需进一步厘清IRF1如何与NF-κB、STAT1等转录因子协同,精确调控保护性免疫与病理性炎症之间的表观遗传平衡。
参考文献:
Ayala J M, Bellworthy R, Mancini M, et al. Pioneer factor IRF1 unlocks latent enhancers to rewire chromatin and immunometabolism in inflammatory macrophages. bioRxiv, 2026: 2026.02. 27.708404.


