
rG4-seq上线,捕获RNA G-四链体折叠图谱
rG4-seq
RNA G-四链体测序
rG4-seq测序技术通过比较Li⁺与K⁺条件下逆转录酶停滞(RT Stalling,RTS)信号差异,在转录组水平实现RNA G-四链体(RNA G-Quadruplex,rG4)结构的精准鉴定与定位。rG4是RNA分子中重要的高级结构元件,在RNA稳定性、可变剪接、翻译调控及疾病发生等过程中发挥不可或缺的调控作用。
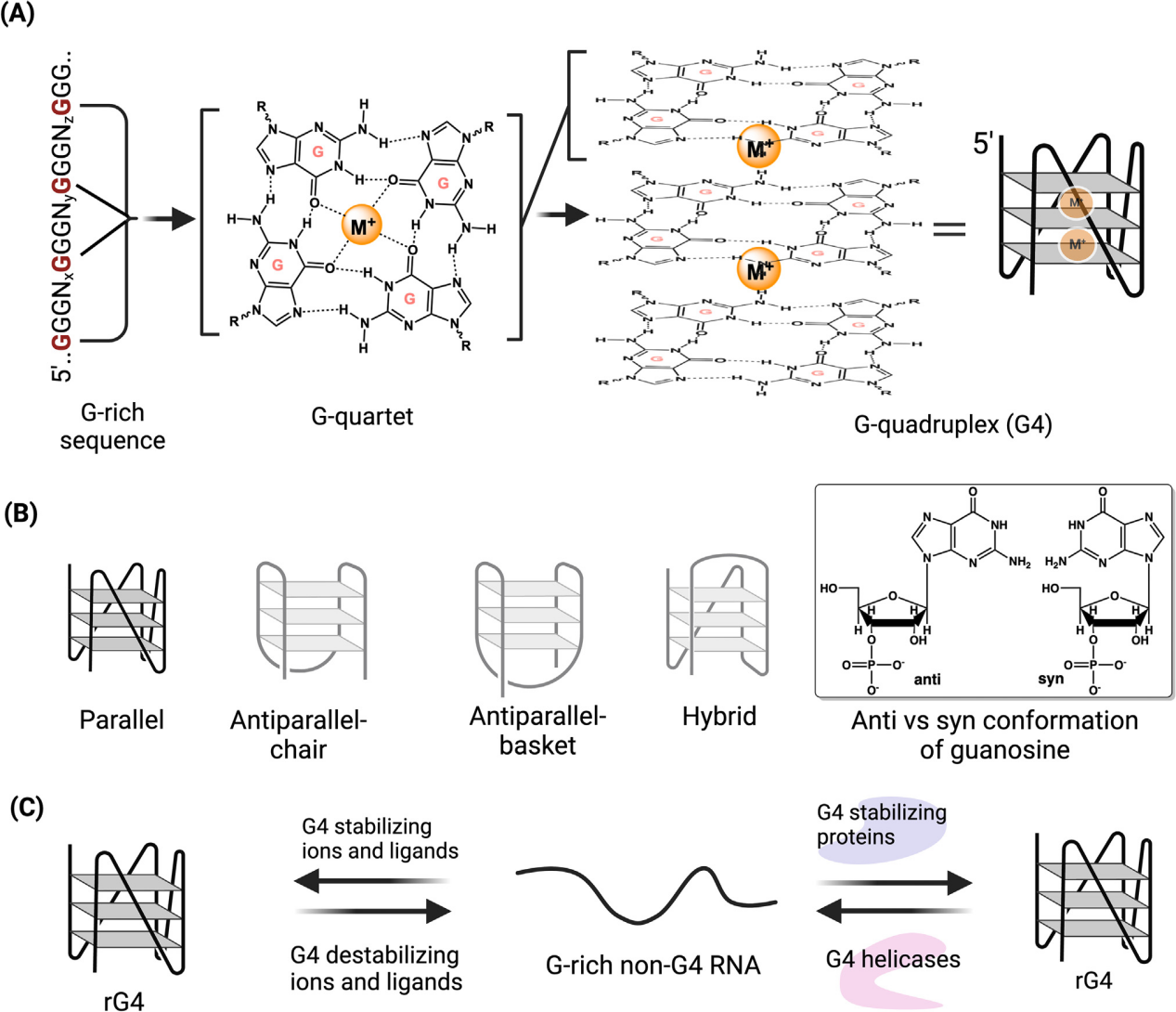
什么是rG4?
rG4是一类由富含G(鸟嘌呤)的RNA序列折叠形成的四链高级结构,其核心由多个G-四聚体(G-quartet)通过Hoogsteen氢键互相连接后堆叠而成,并在中心金属阳离子(特别是K⁺)存在下形成一个坚固的四股螺旋构型。近年来研究发现,rG4广泛存在于mRNA、lncRNA及其他非编码RNA中,可通过调控RNA的转录后加工、可变剪接、亚细胞定位、翻译效率和稳定性等过程影响基因表达。rG4异常形成或调控失衡与肿瘤、神经退行性疾病、病毒感染等多种疾病密切相关,因此已成为RNA结构生物学和RNA药物研发领域的重要研究热点之一。
rG4已知的关键功能作用
翻译调控是rG4最核心的功能,位于5' UTR的rG4通过阻碍核糖体扫描抑制翻译起始,而位于ORF内的rG4可导致核糖体停滞,从而抑制翻译延伸;部分rG4也可作为内部核糖体进入位点(IRES)的组成元件促进翻译【2-4】。
在前体mRNA加工方面,rG4参与可变剪接位点的选择(如BCL-x、FMRP相关转录本),并通过影响3' UTR中的poly(A)信号位点选择调控选择性多聚腺苷酸化,进而产生不同长度的转录本【2,5,6】。
在mRNA稳定性与定位方面,细胞应激(氧化应激、饥饿)可诱导3' UTR rG4广泛折叠,显著增强mRNA稳定性,且该过程在应激解除后可逆;rG4也可作为神经元mRNA树突靶向运输的定位信号【7,8】。
在miRNA调控方面,3' UTR rG4可干扰miRNA与靶位点的结合,而pri-miRNA及pre-miRNA中的rG4结构影响Drosha和Dicer的加工效率,从而调控miRNA的生物发生【5,9】。在端粒维持与端粒酶调控方面,端粒重复序列RNA(TERRA)中的rG4与TRF2的GAR结构域相互作用以维持端粒稳定性,而端粒酶RNA组分(TERC)中的rG4可干扰P1螺旋的形成,进而调控端粒酶活性【5,10】。
在相分离与无膜细胞器方面,rG4通过与含RGG结构域的RNA结合蛋白协同作用,促进液-液相分离,参与应激颗粒、P-bodies及旁斑(paraspeckles)等无膜细胞器的形成与调控【3,5】。
此外,rG4还参与线粒体基因表达调控(GRSF1解折叠线粒体RNA中的rG4以促进线粒体mRNA表达),以及病毒免疫逃逸(EBNA1、LANA等病毒mRNA中的rG4抑制病毒蛋白翻译并调控抗原呈递)【3】。尤其是,rG4的折叠状态受RNA解旋酶(如DHX36)及m6A等RNA修饰的动态调控,其功能的失调与神经退行性疾病、肿瘤等多种人类疾病密切相关【2-4】。
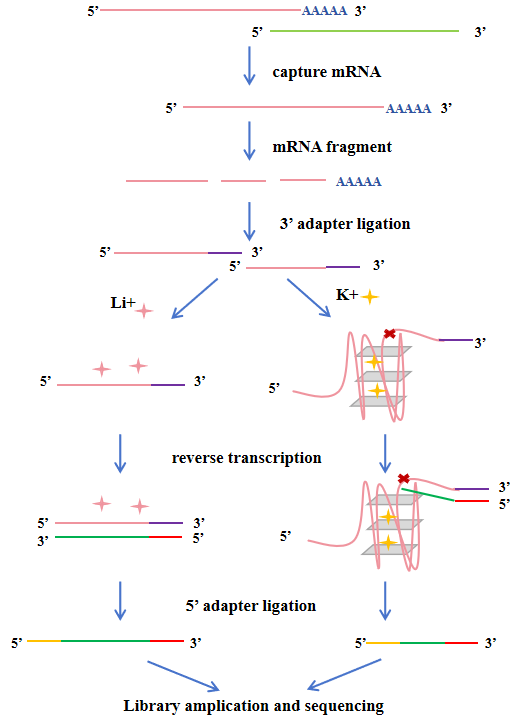
Li+和K+各2个重复,一个样4个文库,需要另外设置生物重复
技术流程
rG4-seq的优化升级
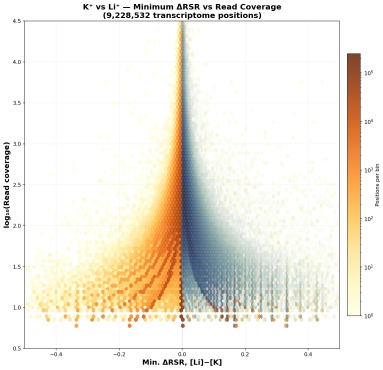
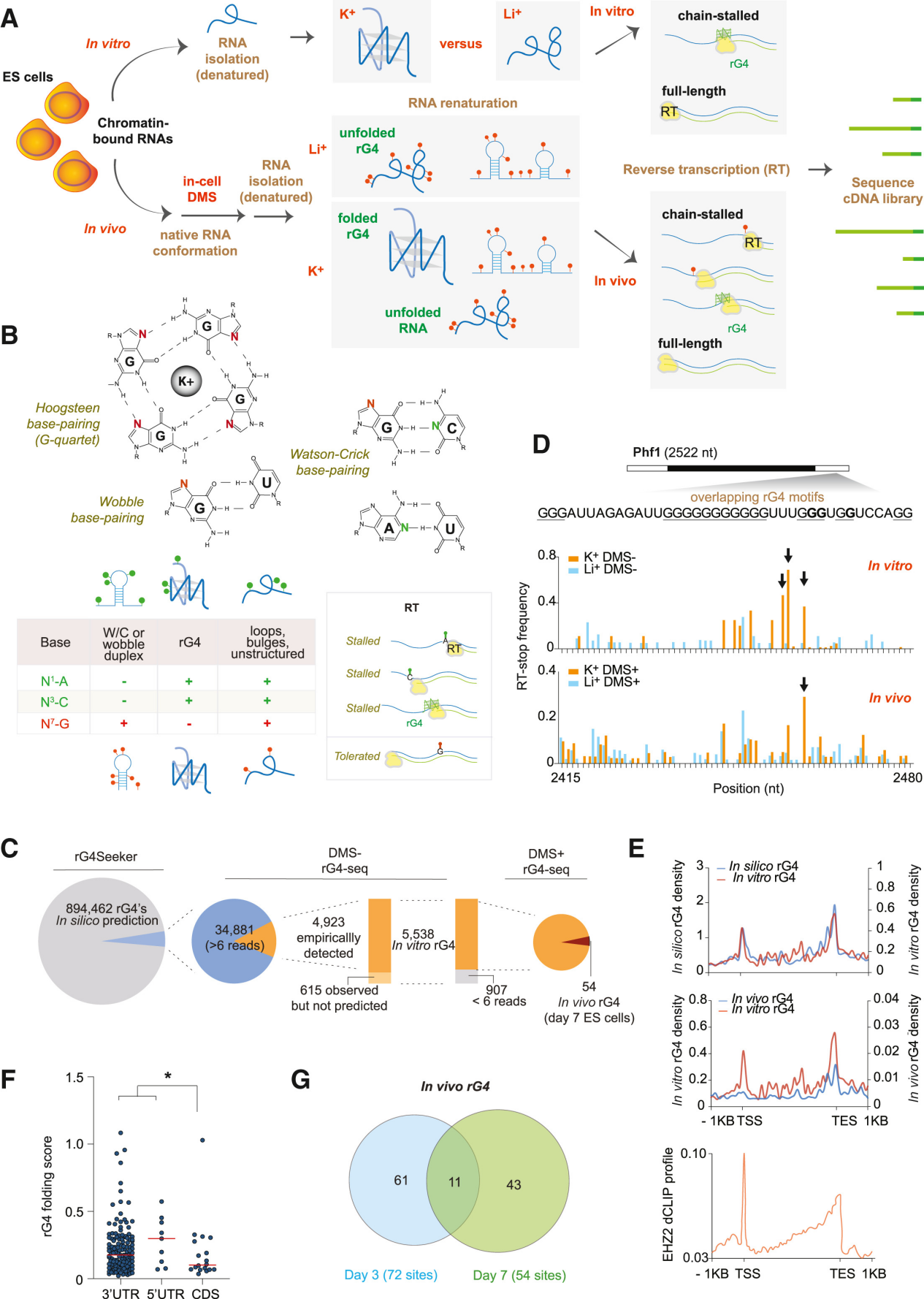
当逆转录酶沿着RNA模板合成cDNA时,如果遇到已形成的rG4结构,会因其空间位阻而发生特异性停滞。这种停滞会导致cDNA合成提前终止,其断点恰好位于rG4结构的3'端。rG4-seq通过三种不同的条件来区分由rG4引起的RTS与背景噪音,包括Li+(背景对照,解链rG4)、K⁺(生理条件,稳定rG4)、K⁺ + PDS(配体条件,强效稳定rG4)。
在rG4稳定条件(K⁺或K⁺+PDS)下,由于逆转录在rG4处停滞,测序reads在rG4位置的覆盖度会出现明显下降;而在对照条件(Li⁺)下则无此现象。通过计算并比较K⁺或K⁺+PDS组相较于Li+对照组的覆盖度骤降(即停滞)情况,利用线性模型拟合和统计学检验(FDR ≤ 0.1),筛选出高置信度的RTS位点,从而在全转录组水平确定经典的和非经典的rG4结构【11】。有研究团队针对rG4-seq的技术流程和分析方法进行了优化,开发了rG4-seeker【12】,能有效消除背景噪音,过滤假阳性信号,不再需要大量重复。
原方案是三种条件(L⁺、K⁺、K++PDS)各4个重复(4×3),共12个文库。而rG4-seeker开发了不依赖重复的分析工作流,结果表明两个重复的rG4-seq实验足以消除噪音,并且K⁺+PDS条件并非必需,意味着可缩减为仅2个条件(L⁺和K⁺)和2个重复(2×2)。在保证高置信度的同时,降低检测成本和确保准确性。
送样要求
仅限人、大小鼠,其他物种需评估
≥1×10^7个细胞,加TRIzol裂解液,干冰寄送
活细胞样本
≥200 mg,加 TRIzol裂解液,干冰寄送
组织样本
总量≥50 μg;RIN≥6,确保RNA无降解
Total RNA
数据分析
分析内容
1. 数据质控
2. 比对基因组
3. rG4-四链体检测
4. rG4结构类型注释
5. rG4基因组区域分布
6. rG4位点top基因排布
7. rG4位点差异分析
8. 差异rG4位点富集分析
rG4位点Li+和K+条件分布
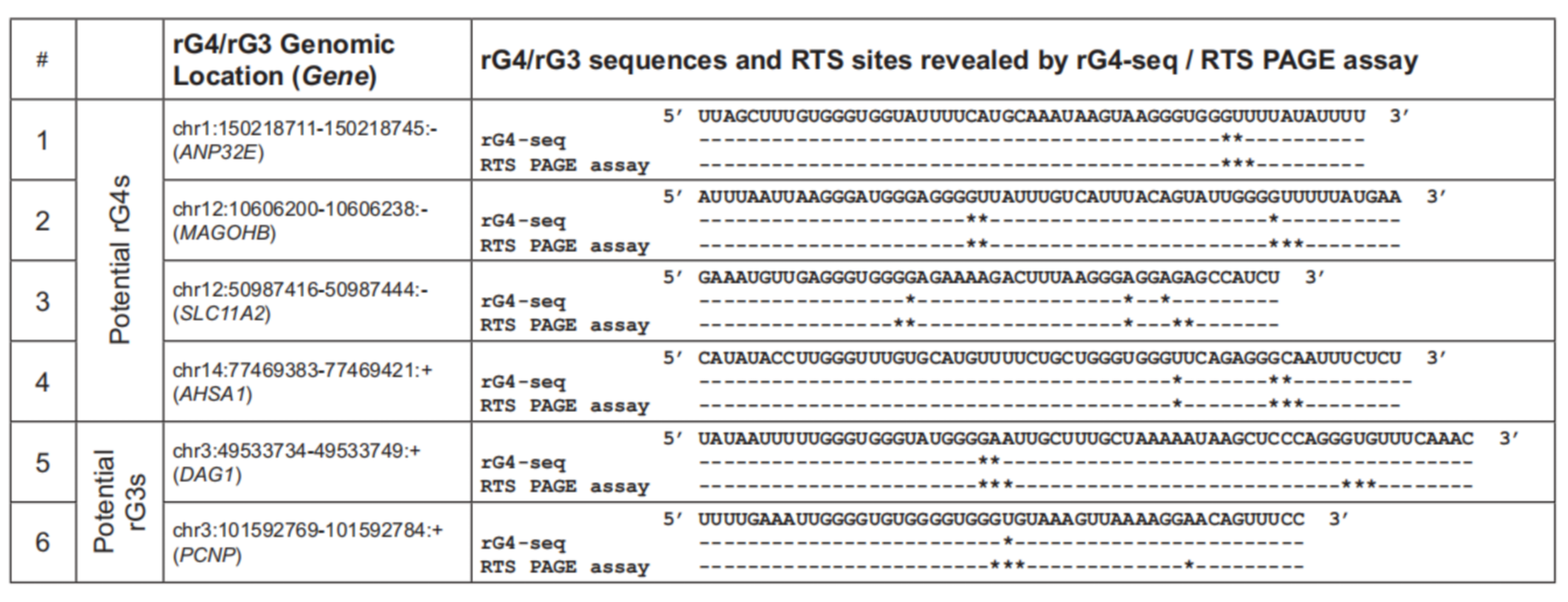
rG4位点展示
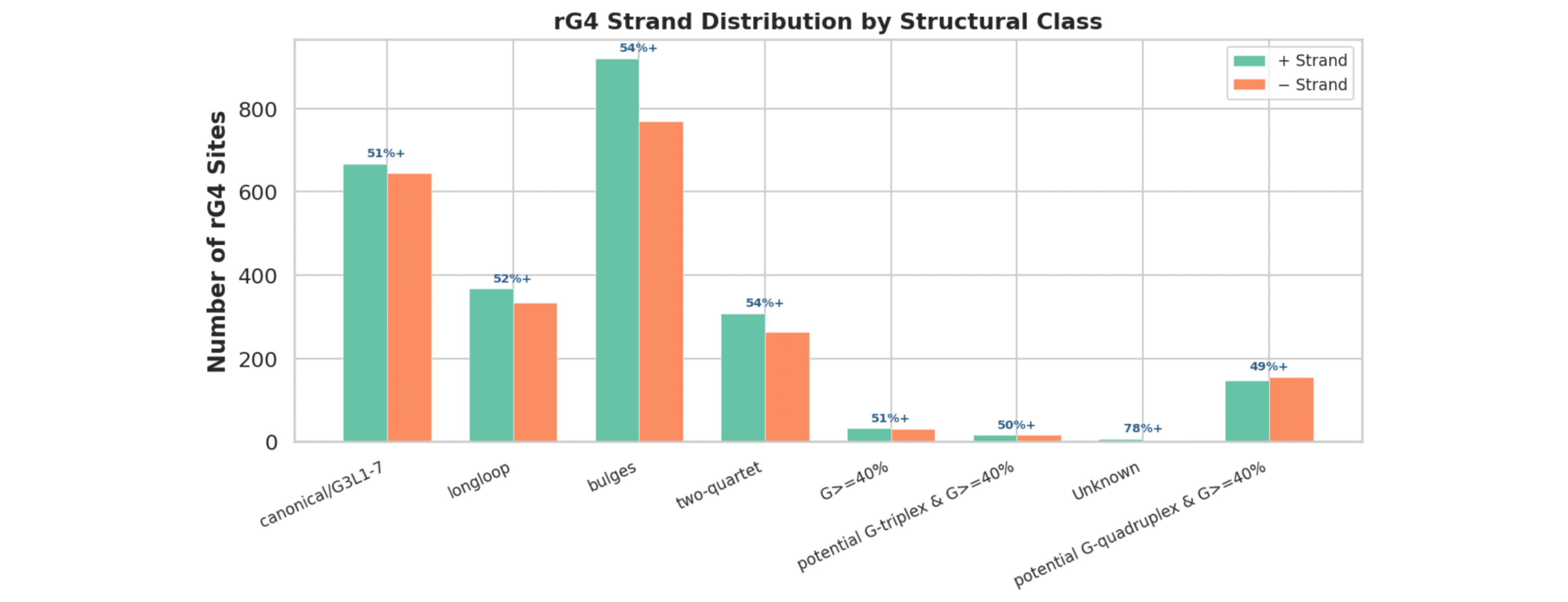
rG4位点正负链分布
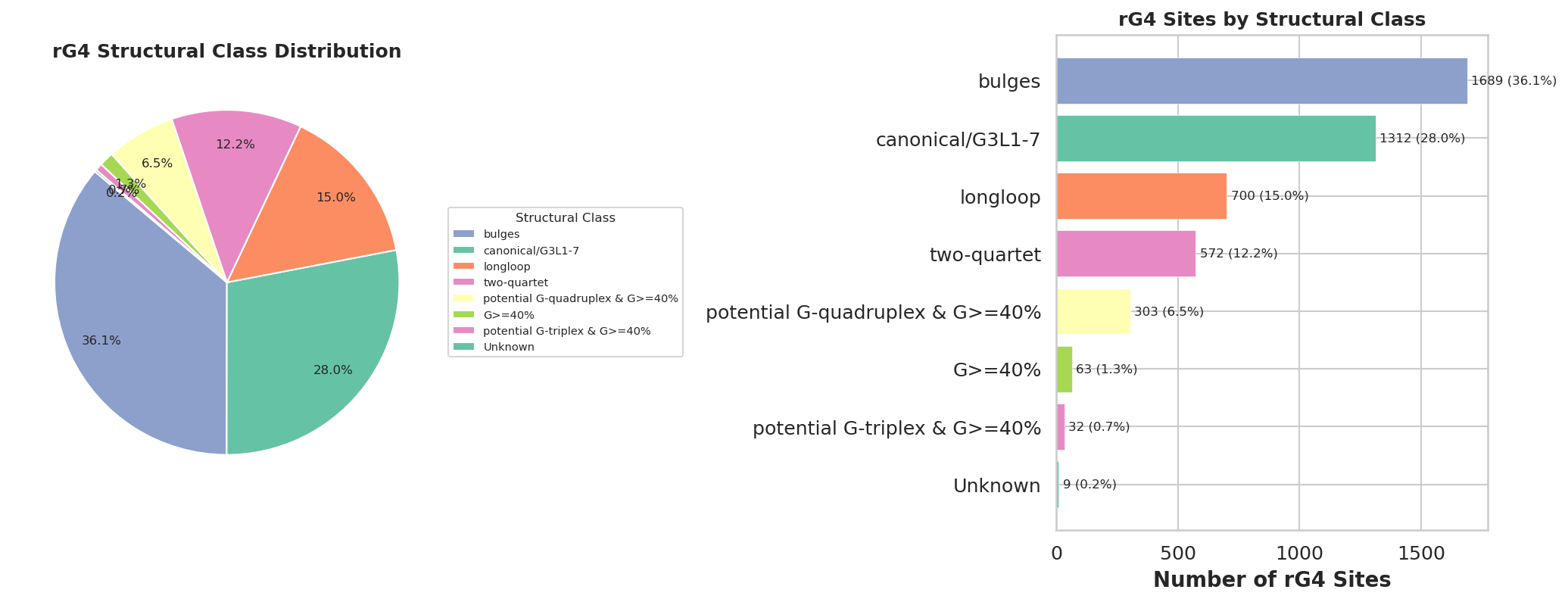
rG4位点注释
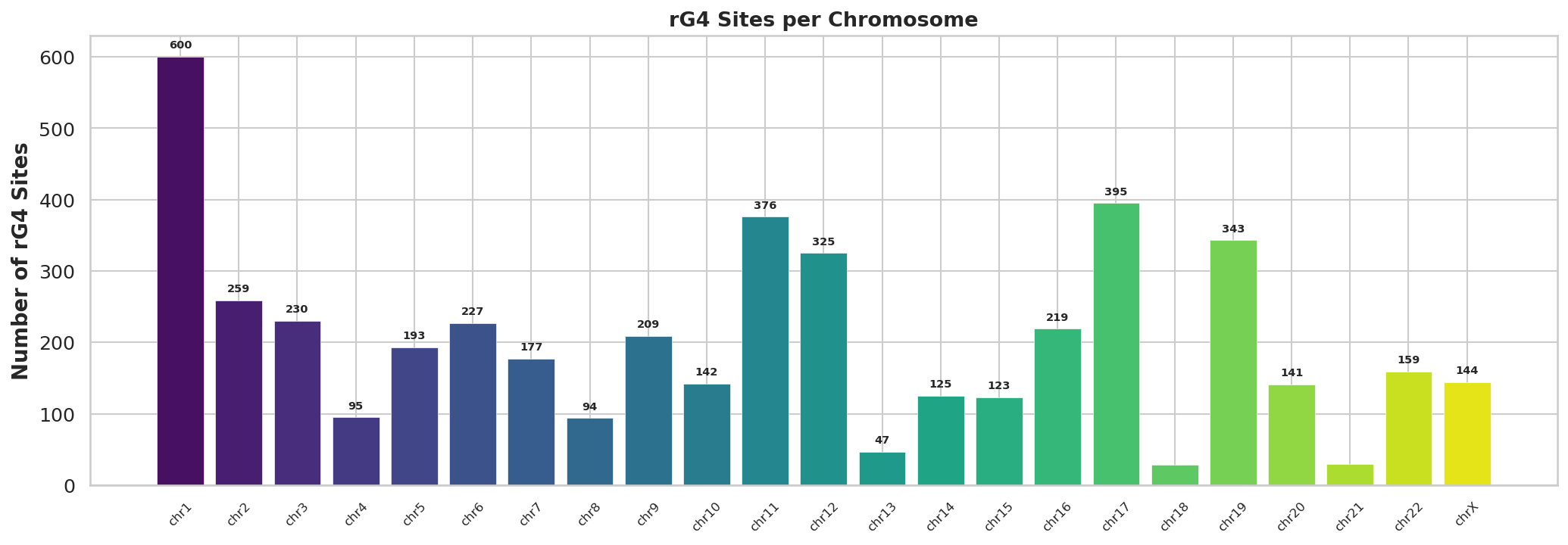
rG4位点基因组分布
rG4-seq研究案例
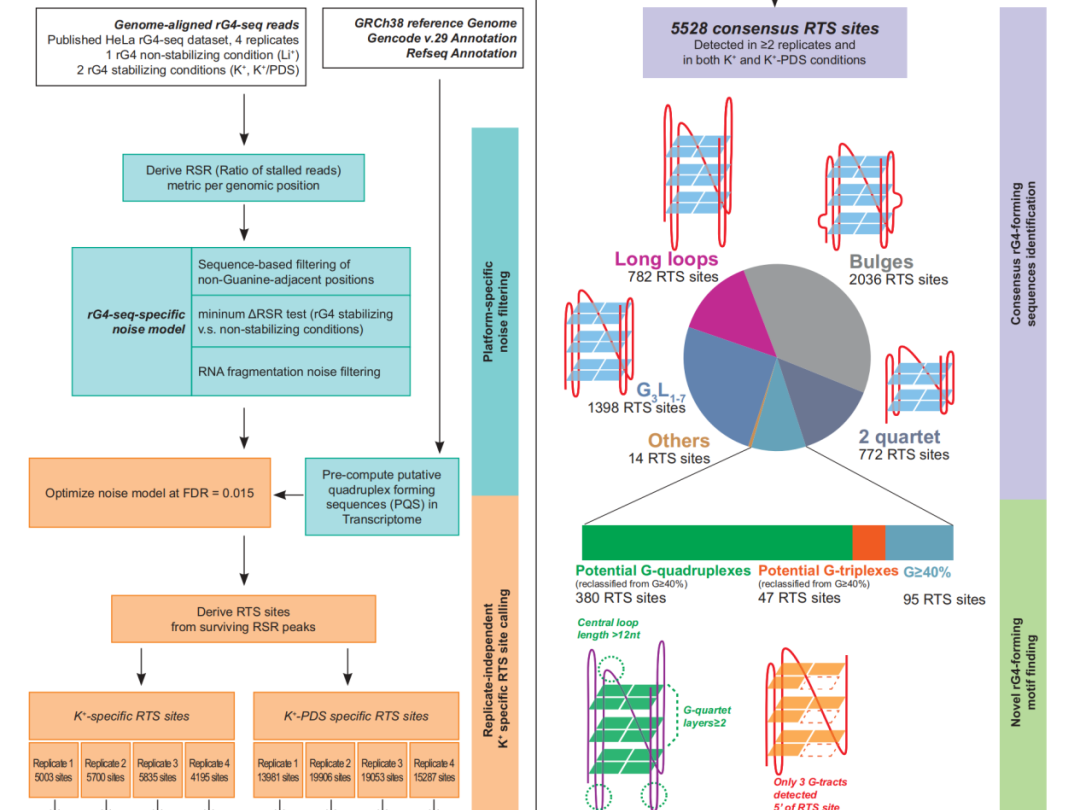
1、rG4-seeker enables high-confidence identification of novel and non-canonical rG4 motifs from rG4-seq experiments【12】
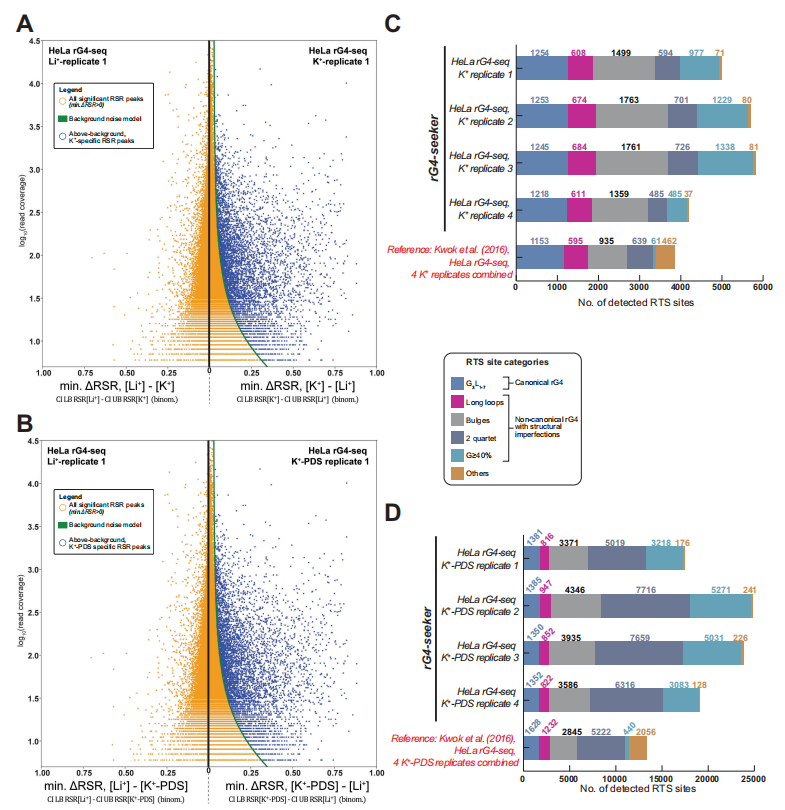
作者开发了rG4-seeker,并集成于rG4-seq数据集进行分析,从转录组层面共识鉴定出5528个高置信度RTS位点,假阳性(Others类)仅14个,相比原方法在降低假阳性的同时显著提升了非经典rG4(Bulges、2-quartet等)的检出数量。

在新型结构发现方面,额外鉴定出380个新型潜在rG4和47个潜在RNA G-三联体(rG3),并通过PAGE-based RTS体外实验逐一验证,所有6个候选位点均得到证实。分析结果还揭示了一条此前未知的规律:rG4对结构缺陷的容忍度呈5′→3′递减趋势,即5′端的G-tract可更短、连接环可更长,而3′端则相反。
另外,作者利用rG4-seeker的重复独立特性系统评估了重复数量与检测重现性的关系,发现两重复实验可重现约70%的RTS位点,建议将实验规模从12个文库精简至4–6个文库,显著降低实验成本。
2、Differential Evolution of CDS and UTR Non-canonical RNA G-quadruplex Structures in Eukaryotic Transcriptomes【13】
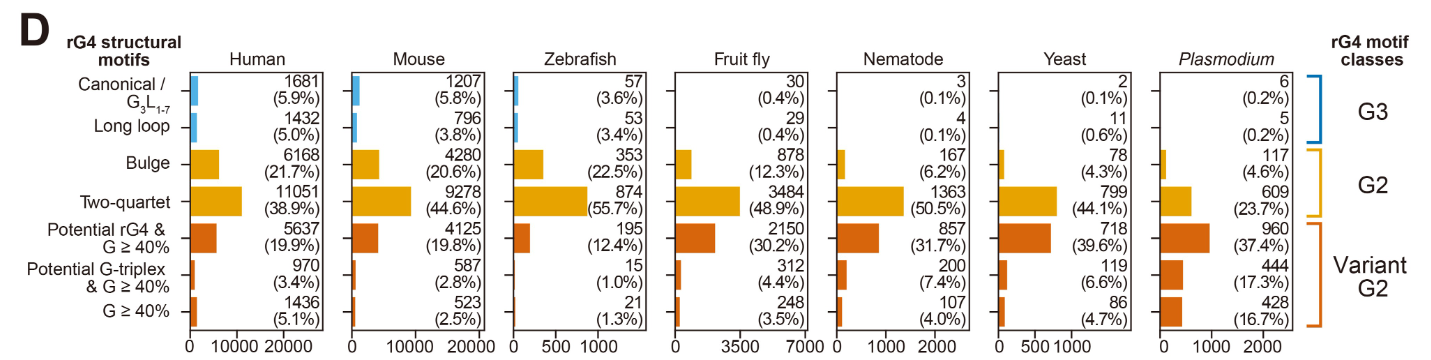
该研究利用rG4-seq技术对7个真核模式生物的转录组进行了全面的rG4结构图谱绘制。通过在Li⁺、K⁺及K⁺/PDS三种缓冲条件下捕获RTS,作者系统鉴定了各物种中的rG4结构,发现非经典rG4(尤其是G2 motif)在所有物种中均占主导地位,且rG4组的规模随进化复杂度整体扩张。
在CDS rG4的分析中,作者发现大多数CDS rG4与重复元件或低复杂度区域无关,其G-tract组成与密码子偏好密切相关——原始真核生物以G为主,而高等真核生物中谷氨酸(Glu)和缬氨酸(Val)的使用显著增加,揭示了CDS rG4随氨基酸偏好演变而逐步积累的进化模式。
在基因家族层面,作者发现古老基因家族(Eukaryota、Bilateria、Vertebrata 进化支)及脊椎动物全基因组加倍保留的ohnolog基因更倾向于携带rG4,支持了"rG4 在基因诞生后才被插入转录组"的假说。
3、G-quadruplex folding in Xist RNA antagonizes PRC2 activity for stepwise regulation of X chromosome inactivation【14】
该文章同样使用rG4测序技术系统绘制了小鼠ESC分化过程中rG4的体内折叠图谱。结果发现,转录组中虽有近90万个潜在rG4基序,但体内真正稳定折叠的仅有54个,且这些折叠位点在分化不同阶段(Day 0/3/7)呈现高度动态变化、时间特异性强。在Xist RNA上,作者鉴定出AF、E7和3'三个rG4簇,其中3' rG4在分化期(Day 7)折叠最为稳定。生化实验证明,折叠的rG4能以高亲和力结合PRC2(Kd低至46 nM),并强效抑制其组蛋白甲基转移酶活性(IC₅₀约57–69 nM)。进一步通过CRISPR敲除3' rG4后,PRC2失去"刹车"而过度激活,导致Xi染色体上H3K27me3大量超富集,但Xist-PRC2复合体同时被困于S1区室、无法向S2转移,最终造成基因沉默失败。使用rG4稳定剂处理细胞得到完全相反的表型,进一步验证了上述结论。据此,作者提出rG4折叠作为"分子变阻器"调控PRC2的"Hit-and-Run"模型,揭示了RNA结构动态调控染色体表观遗传沉默的新机制。
参考文献
1.Kharel P, Ivanov P. RNA G-quadruplexes and stress: emerging mechanisms and functions. Trends Cell Biol. 2024;34(9):771-784.
2.Lyu K, Chow EY, Mou X, Chan TF, Kwok CK. RNA G-quadruplexes (rG4s): genomics and biological functions. Nucleic Acids Res. 2021;49(10):5426-5450.
3.Dumas L, Herviou P, Dassi E, Cammas A, Millevoi S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem Sci. 2021;46(4):270-283.
4.Kharel P, Becker G, Tsvetkov V, Ivanov P. Properties and biological impact of RNA G-quadruplexes: from order to turmoil and back. Nucleic Acids Res. 2020;48(22):12534-12555.
5.Li XD, Yi C, Yin HH, Zhang H. New advisory board members speak on challenges and opportunities in chemical biology. Cell Chem Biol. 2023;30(11):1324-1326.
6.Beaudoin JD, Perreault JP. Exploring mRNA 3'-UTR G-quadruplexes: evidence of roles in both alternative polyadenylation and mRNA shortening. Nucleic Acids Res. 2013;41(11):5898-5911.
7.Kharel P, Fay M, Manasova EV, et al. Stress promotes RNA G-quadruplex folding in human cells. Nat Commun. 2023;14(1):205. Published 2023 Jan 13.
8.Subramanian M, Rage F, Tabet R, Flatter E, Mandel JL, Moine H. G-quadruplex RNA structure as a signal for neurite mRNA targeting. EMBO Rep. 2011;12(7):697-704. Published 2011 Jul 1.
9.Rouleau S, Glouzon JS, Brumwell A, Bisaillon M, Perreault JP. 3' UTR G-quadruplexes regulate miRNA binding. RNA. 2017;23(8):1172-1179.
10.Varshney D, Spiegel J, Zyner K, Tannahill D, Balasubramanian S. The regulation and functions of DNA and RNA G-quadruplexes. Nat Rev Mol Cell Biol. 2020;21(8):459-474.
11.Kwok CK, Marsico G, Sahakyan AB, Chambers VS, Balasubramanian S. rG4-seq reveals widespread formation of G-quadruplex structures in the human transcriptome. Nat Methods. 2016;13(10):841-844.
12.Chow EY, Lyu K, Kwok CK, Chan TF. rG4-seeker enables high-confidence identification of novel and non-canonical rG4 motifs from rG4-seq experiments. RNA Biol. 2020;17(7):903-917.
13.Chow EY, Zhao J, Kwok CK, Chan TF. Differential Evolution of CDS and UTR Non-canonical RNA G-quadruplex Structures in Eukaryotic Transcriptomes. Genomics Proteomics Bioinformatics. 2025;23(6):qzaf078.
14.Lee YW, Weissbein U, Blum R, Lee JT. G-quadruplex folding in Xist RNA antagonizes PRC2 activity for stepwise regulation of X chromosome inactivation. Mol Cell. 2024;84(10):1870-1885.e9.